TYPE: Research Article ![]()
Tushar Parab¹ , Salvador Lyngdoh¹*
, Salvador Lyngdoh¹*
¹Dept. of Landscape Level Planning and Management, Wildlife Institute of India, Chandrabani, Dehradun, 248001.

Introduction
Among terrestrial animals, carnivores are the most threatened groups (Karanth et al., 2009). Smaller carnivores may be helpful indicator species in preserving ecosystems since their specialization and resource selectivity are typically stronger than those of large carnivores. Given that these carnivores compete on numerous fronts when there are significant nutritional overlaps across species and intermediate disparities in body size, interspecific killing occurs more often (Donadio & Buskirk, 2006). Due to this threat, they must compromise (Laundre et al., 2010) to survive in a challenging environment and avoid dominant predators (Bischof et al., 2014). Additionally, because ecological niches overlap, interspecific competition is considered one of the most significant mechanisms limiting the number of species that may exist within an assemblage (Di Bitetti et al., 2010). Hence, competitive interactions can shape patterns of community assemblages, determine how niches are divided, and support the development of coexistence strategies among competing species (Chiang et al., 2012; Cristescu et al., 2013). Interspecific competition determines sympatric species’ demographic and ecological traits with effects of utilization and interference (Grassel et al., 2015).
Knowledge of South Asian wild cats is limited due to their elusive behavior (Nowell & Jackson, 1996.). The leopard cat ( Prionailurus bengalensis) is a small, widely distributed species in Asia that inhabits a broad range of natural habitats, particularly in areas where humans have altered the landscape (Marcella, 2004).
The International Union for Conservation of Nature has categorized it as a species of “least concern” since it is not currently considered to be threatened (https://www.iucnredlist.org) Most density estimates for smaller cats are typically derived from radiotelemetry studies using a small sample size. ( e.g., Dillon & Kelly, 2008; Dunstone et al., 2002 ; Sliwa, 2004). Only a few studies have used camera trapping data and capture and recapture models to estimate densities of smaller cat species, and the majority of those studies are limited to smaller data sets and are mainly restricted to the Americas ( e.g., Reppucci et al., 2011; Trolle & Kery, 2003) and Europe (Kéry et al., 2011- European wild cat Felis silvestris; based on DNA) and are merely available for smaller cat species of Asia and Africa.
Across its entire range, the red fox ( Vulpes vulpes) is well known for its adaptive and opportunistic behavior (Macdonald, 1983). It is one of the most studied carnivore species, and it inhabits a variety of habitats, from the Arctic to subtropical regions (Nowak, 1999). This resilient mesocarnivore inhabits both human-dominated and natural environments over various geographic areas (Bidlack et al., 2006; Macdonald & Reynolds, 2008). The Yellow-Throated Marten, being susceptible to habitat degradation and disturbance, represents an indicator species for the forest ecosystem (Harrison et al., 2004; Taylor & Buskirk, 1994). The diet of this carnivorous generalist ranges from frugivorous to hyper-carnivorous. The yellow-throated marten has an omnivorous diet and is a semi-arboreal, meso- carnivorous species (Choi et al., 2015; Nandini & Karthik, 2007). Key ecological characteristics, including size, home range and geographical distribution of our three study species, are provided in Table 1. The leopard cat is a strongly nocturnal species, while the yellow-throated marten is typically diurnal (Bhatt et al., 2021).
Two key ecological processes are the abundance dynamics of these small carnivores, and the temporal and spatial variations in their habitat use. Several studies on carnivores have revealed that sympatric species typically exhibit diverse patterns of activity and habitat utilization (Seidensticker, 1976; Bothma et al., 1984; Sunquist & Sunquist, 1989; Grassman et al., 2005), depending on the possibilities for habitat partitioning offered by heterogeneous landscapes (Lantschner et al., 2012). How do the behaviors exhibited by leopard cats (Prionailurus bengalensis) towards red foxes ( Vulpes vulpes) and yellow-throated martens ( Martes flavigula), as evidenced by occupancy, density, abundance estimates, and spatial/ temporal interactions, inform our understanding of resource partitioning and competitive dynamics within their shared habitat? Here, using occurrence data from camera traps, we examine coexistence patterns among a guild of two small sympatric Himalayan carnivores (red fox and yellow- throated marten) with leopard cats, which together form an ecological guild inhabiting forested areas of the Western Himalayas.
Materials and Methods
Study Area:
This study was conducted in Dhauladhar Wildlife Sanctuary (982.8 km²), which is located at the junction of the 2C- Western, 1B-North West, and 1A-Trans Himalayan ranges (Rodgers & Panwar, 1988) in Himachal Pradesh. Because of the significant variance in altitudinal zoning, the habitats are diverse and host a variety of species. For six months (June to November 2021), camera traps were used to collect evidence of the animal presence to analyze the diversity, density, abundance, occupancy, and activity patterns. The locations of the camera traps (n = 35) were selected based on the presence of carnivore signs, access, terrain features, and animal trails (Figure 1). Camera trap locations were set in 2 km × 2 km grid cells (n = 178), out of which 35 grids were sampled, falling in accessible locations within the sanctuary boundaries. As this study was part of a broader project on mammalian diversity, the grid size (4 km²) was chosen to represent a home range of an intermediate size between that of the smallest (leopard cat) and largest (Himalayan brown bear) carnivores in the study area and to maximize the spatial coverage. In addition, 4 km² sampling grids sufficiently cover the home range of our study species (Table 1), while providing significant spatial coverage to capture variability within and between home ranges. Each grid contained an individual camera trap (Cuddeback color C2), active for 30–35 days. The camera model and type were the same across all stations, and cameras were placed 3 meters from the closest track with a standardized lens height of 40 centimeters. The sum of all camera days within the sample period was used to define the total amount of trapping effort (1,295 days).
Table 1: Ecological characteristics of three study species found in Dhauladhar Wildlife Sanctuary, Himachal Pradesh, India: leopard cat (Prionailurus bengalensis), red fox (Vulpes vulpes), and yellow-throated marten (Martes flavigula).
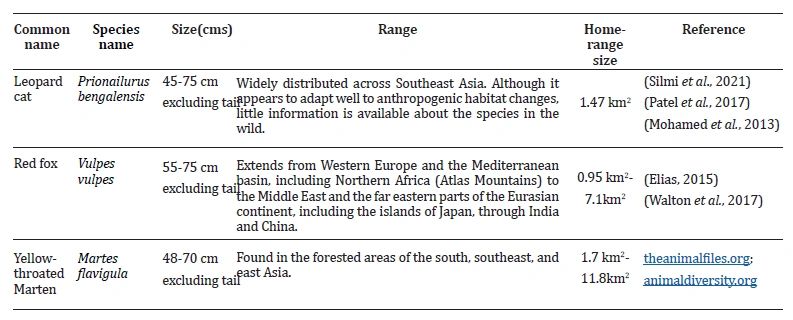
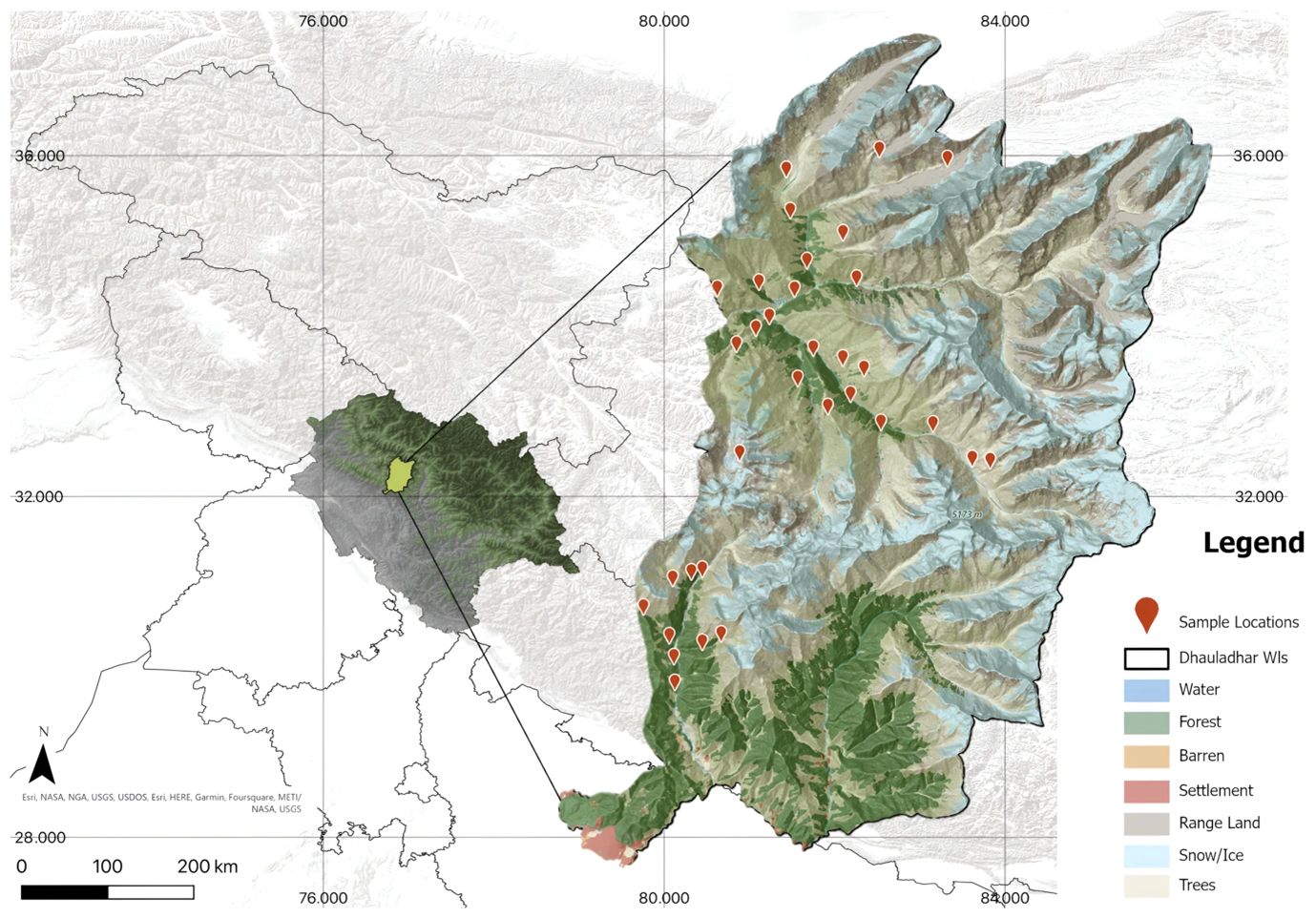
Figure 1: Locations of camera trap stations spaced throughout Dhauladhar Wildlife Sanctuary, overlaid on land cover (Source: ESRI, USGS 2022).
Individual identification and abundance model for spatially replicated counts:
As the number of photo captures of leopard cats was higher with left flanks, we considered it to estimate the density. The markings on the pelage of the rear limbs, forelimbs, and forequarters were visually inspected to identify individual leopard cats from images. We used morphological types and features like tail, foot, and forearm for individual identification of red fox following (Sarmento et al., 2009), as represented in Figure 2, considering analysis for a single season within a small time frame with limited captures (n=105). Similarly, individual identification of leopard cats was also based on morphological traits like spot patterns (Figure 3). While individual identification is generally used in SECR models to estimate animal populations, yellow-throated martens could not be included in these analyses due to the difficulty in differentiating between individuals based on coat patterns. Therefore, yellow-throated martens, which need individual identification for precise density estimations, were excluded from the SECR analysis. Instead, we used camera trap distance sampling to estimate the density of yellow-throated martens. With this method’s help, we overcame the limitations of individual identification and achieved reliable density estimates for this species.
It is essential to interpret relative abundance indices from different sampling techniques to estimate population size correctly. N-mixture models were applied by the pcount function in the ‘unmarked’ package (Fiske & Chandler, 2011) in R (R Core Team, 2019) software. The hierarchical model approach and repeated count data are used in this program to estimate abundance, and the maximum likelihood method is used to fit the models. However, the results of the abundance models were interpreted only as a change in species abundance with respect to covariates and not to estimate the absolute population size following (Gilbert et al., 2021). Based on a review of the literature (Table 2), we selected ecologically significant variables and checked if they were correlated (Supplement 1). Highly correlated habitat covariates (>0.7) were excluded. All layers were prepared using Quantum GIS Tisler version 3.24. We used elevation, slope, aspect, distance to the road and human footprints as predictor variables for leopard cats and elevation, distance to the road, human footprints, and landcover type for red foxes. Using this N-mixture abundance model, our main objective was to investigate how the regional abundance of red foxes affects the regional abundance of a subordinate taxon, the leopard cat.
Spatially explicit capture-recapture density estimation.:
SECR models are based on the fact that animals are more likely to be detected if they spend more time close to detectors. The SECR model needs two parameters: (1) detection probability (g0) and (2) spatial scale at which encounter rates or detection probabilities decrease when animals’ activity centers and detectors are located further apart (σ). Although alternative relationship forms can be considered, the form is often assumed to be half-normal (Howe et al., 2022). Model assumptions were that 1: animals had stable activity centers ( i.e., there was no ongoing emigration or immigration) and classified captures as binary events (‘proximity’ as the detector type); and 2: we assumed that the detection process was a continuous function through the activity centers to the point of no capture probability, and modeled detection as a half-normal function. (Borchers et al., 2008). We used package secr (Efford, 2012) in R to evaluate density estimates of leopard cats and red foxes in the study area. For the leopard cat and red fox, the trap array
Table 2: Landcover, topographic, and anthropogenic covariates used to model abundance, densities, and occupancy of leopard cat, red fox, and yellow-throated marten in Dhauladhar wildlife sanctuary.
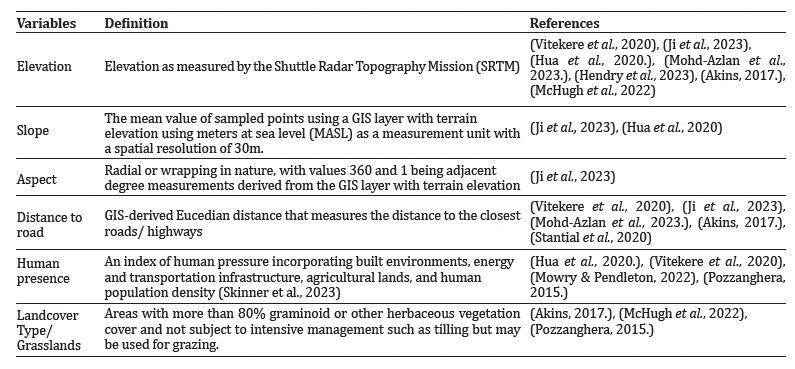

Figure 2: Individual identification of red fox for generating count data used in N-mixture modeling and SECR. Two individuals, A – B, C – D, and E – F, were photo-captured at different camera trap locations during one sampling occasion.
buffer used was 8,000m and 12,000 m, respectively, which was four times the value of sigma, so that any individuals with activity centers outside the trap array could be included in the density estimation.
Space-use patterns: multispecies occupancy model:
While accounting for imperfect detections, a multispecies occupancy model (Rota et al., 2016) was used to access the spatial interactions of leopard cats, red foxes, and yellow-

Figure 3: Individual identification of leopard cat for generating count data used in N-mixture modeling and SECR. Two individuals, A – B, C – D, and E – F, were photo-captured at different camera trap locations during one sampling occasion.
throated marten. A multispecies occupancy model’s fundamental sampling methodology and assumptions are the same as those in the single-species scenario (MacKenzie et al., 2017). We arranged the data obtained from camera traps by creating a detection history into a matrix of sites i by sampled occasions j, selecting one week as a threshold for convergence of models and accuracy of estimates. We employed a two-step process since this model calculates species occurrences individually and co-occurrence factors as an expression of the provided variables (Salvatori et al., 2021). After we chose the covariates that were the most strongly supported for each species, the best variables from the single-species occupancy model were used as predictors of the presence and detection of species individually in the multispecies model. We determined the best models by comparing the Akaike information criterion corrected (AICc) and Akaike weight (AICc weight).
Spatial and Temporal Overlap:
We recorded the date and time of capture visible on the photograph for every image in a dataset. Records of the same species occurring at the same camera trap site within 30 minutes were eliminated to maintain statistical independence and avoid pseudo-replicates. The R package overlap (Meredith & Ridout, 2014) was used to analyze the activity patterns of all three carnivore species and estimate the temporal overlap value. This numerical value ranges from 0 (no overlaps) to 1 (identical activity patterns), calculated using kernel density estimation. Based on the conventional Pianka’s index, which ranges from 0 (no overlap) to 1 (complete overlap), an estimation of spatial niche overlap among the most common species was carried out. Pianka’s niche overlap index was calculated using the Spaa package or Species Association Analysis package (Zhang, 2016) in R Studio.
Results
Abundance and Density Estimates:
We included natural and anthropogenic factors in our co- abundance models as we hypothesized that the abundance of these species would differ in their natural habitat preferences and tolerance to anthropogenic disturbances. Being a topographically diverse area, ranging from an altitude of 1,800-6,000m, we used different habitat covariates for all three species based on predictors that likely can influence the distribution (Table 2). We further examined collinearity among the variables to avoid multicollinearity of the predictor variables (Spearman correlation coefficient r >0.7). Instead of interpreting our findings in terms of abundance, we initially considered how the abundance varies spatially in relation to the variables. Our best model resulted in β = 2.35 individuals of red fox (SE=0.4) per camera station with a detection probability of 0.0237, which was found more abundant than leopard cat (β =0.530, SE=0.1; det p= 0.0229) and yellow-throated marten (β =1.06, SE=0.4; det p= 0.0433). We only report the estimates based on the top models for each species (Table 3). The abundance distribution maps are provided for each species separately (Figure 4).
Table 3: The estimated abundance (β) and detection probability ( det p) of yellow-throated marten, red fox, and leopard cat determined by N-mixture models.
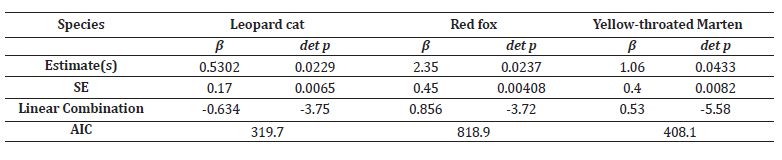
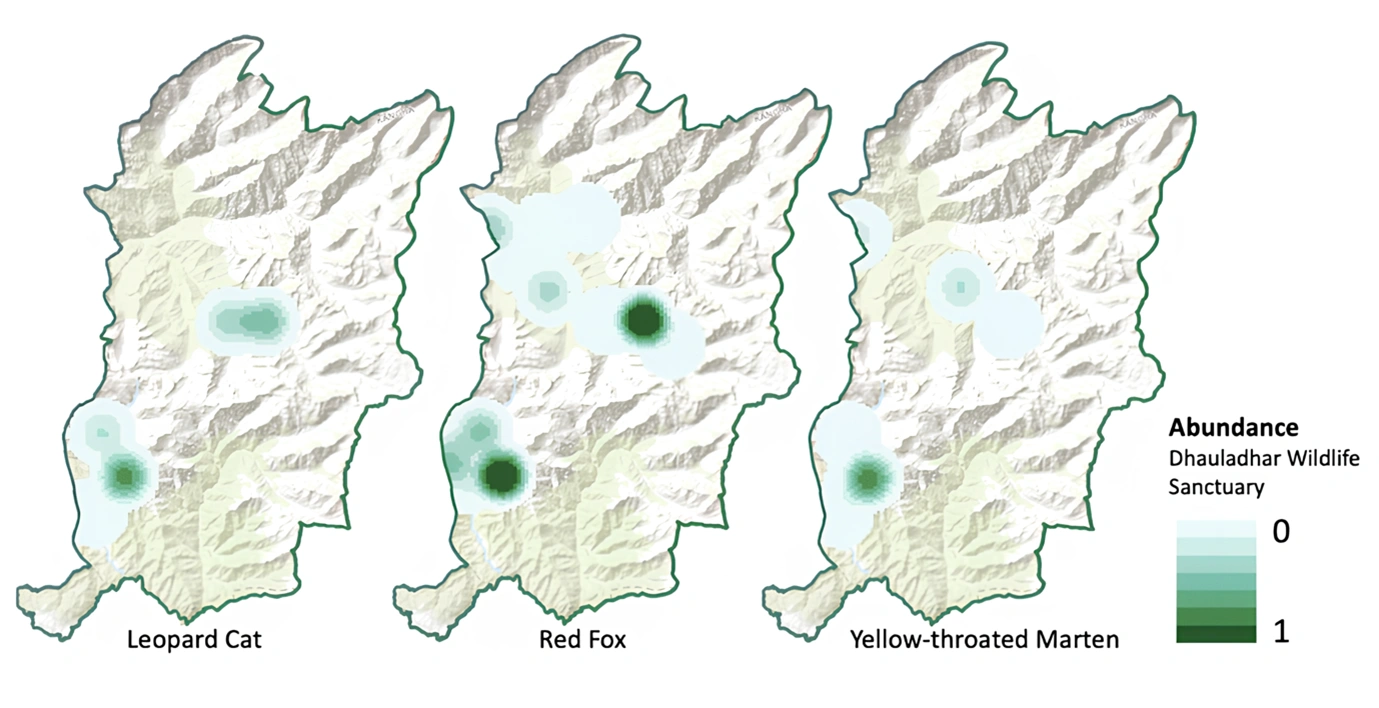
Figure 4: The relative abundance distribution of three study species—leopard cat, red fox, and yellow-throated marten. Value towards 1 represents higher abundance compared to the value towards 0.
The best model (D~1 g0~h2 sigma~1) of a half-normal detection function estimated 18.2 (+-1.38) individuals/100km² for leopard cats (Table 4). The movement of leopard cats between range centers was estimated considering the 296 km² sampling area covered altogether, estimated with an 8000 m buffer around the locations of the camera traps at the periphery (Figure 3). The total number of leopard cats within this area was estimated to be 53.8 individuals. Similarly, for red foxes, our top model (D~1 g0~ h2 sigma~1) of a half-normal detection function predicted 24.6 (+-3.32) individuals/100 km².
Our sampling area was 324 km² based on a buffer of 12,000 m around the outermost camera trap locations, and the overall estimated population from our study region was estimated to be 79.7 individuals. Using camera-trap distance sampling, the density estimates of yellow-throated martens resulted in 44 (+-5.1) individuals/100km². Although this species was excluded from the (SECR) analysis, it remains crucial to know their densities to fully understand the dynamics of the carnivore community in the study area.
Multispecies occupancy model:
In comparison to models that took account of detection covariates, the intercept-only models outperformed. The positive pairwise estimates of the intercept-only models for the leopard cat and red fox (1.929 ± 0.361) suggested that leopard cats were more likely to exist at sites occupied by red foxes (Table 5). The estimates for red fox and yellow-throated marten (0.913 ± 0.40), leopard cat and yellow-throated marten (0.846 ± 0.396) also confirmed co- occurrence of the species in the studied area. The highest overlap in average occupancy (D=0.95, P=0.009) is observed between leopard cats and red foxes (Figure 6), followed by leopard cats and yellow-throated martens(D=0.47, P= 0.039) and red fox and yellow-throated marten (D=0.47, P= 0.039).
Spatial and Temporal Overlap:
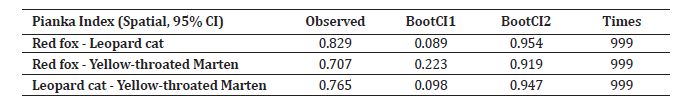
Overall values of Pianka’s indexes of spatial niche overlap were observed the highest between two nocturnal study species (leopard cat and red fox), estimated to be 0.829 with enough sample size, according to comparisons with simulated data. Other two comparisons for nocturnal vs. diurnal species, i.e., leopard cat- yellow-throated marten and red fox- yellow-throated marten, yielded overlap estimates of 0.765 and 0.707, respectively (Table 6). Leopard cat and red fox exhibited nocturnal, while yellow-throated marten exhibited a strictly diurnal activity pattern. Since both leopard cats and red foxes are nocturnal, there was high temporal overlap (D̂4=0.765) and yellow-throated marten being a strictly diurnal species, the estimated temporal overlap of yellow-throated marten and leopard cat was considerably lower (D̂4 =0.120) (Figure 7).
Table 4: Density estimates for leopard cat and red fox using SECR method with maximum likelihood approach in Dhauladhar Wildlife Sanctuary. AICc = Akaike Information Criteria corrected for small sample sizes, g0 = Detection probability, D = Density of individuals per 100 km². The trap buffer for estimating leopard cat and red fox density was 8000m and 12000m, respectively.
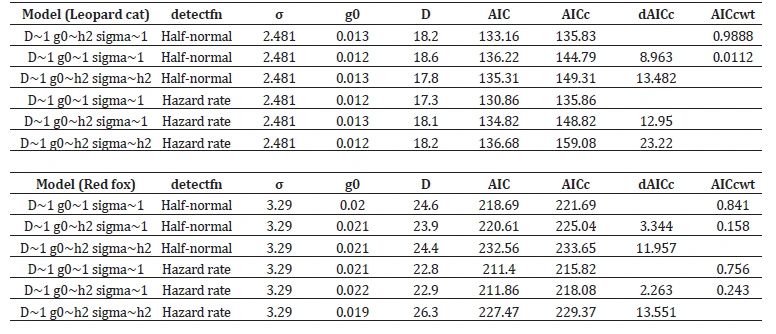
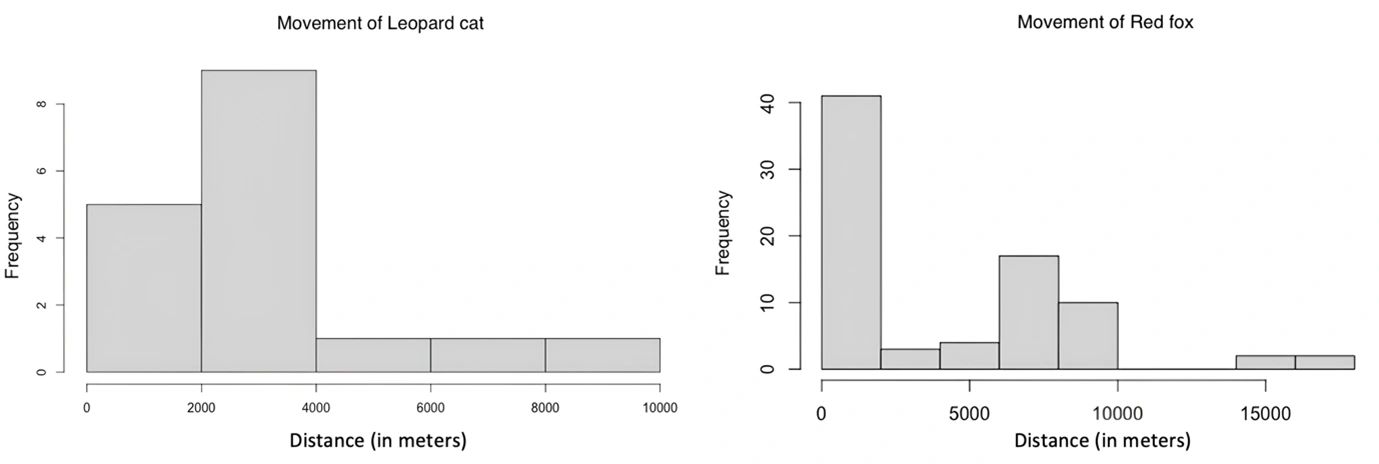
Figure 5: Movement of leopard cats (left) and red foxes (right) between range centers. Using data from the spatially explicit capture-recapture, the plot shows how leopard cats and red foxes move between range centers within the Dhauladhar Wildlife Sanctuary. The distance is represented on the x-axis (in meters), and the frequency of individuals within the designated distance ranges is shown on the y-axis.
Discussion
Due to competitive exclusion, two ecologically similar species could not coexist for an extended period (Macarthur & Levins, 1967). Despite the need for manipulative experiments to conclusively show such depressive competition, our analyses of the relative abundances of red fox and leopard cat show that there is a negative relationship between them, indicating that high red fox (β =2.35) abundance can decrease leopard cat (β =0.53) numbers and could exclude them. Given the inverse relationship between the abundance of red foxes and leopard cats in different habitats, it is possible that leopard cats, the subordinate species, will avoid habitats where red foxes are more prevalent to avoid interference. However, multiple studies at different sites with larger sample sizes are required to establish this interpretation that leopard cats are depressed in habitats where red fox abundance is higher than in habitats where red fox abundance is low. It is essential to understand that the results based on N-mixture modeling provide a competitive link between red fox abundance and leopard cat abundance. Still, on the other hand, results of occupancy modeling suggest that both species are forced to co-occur due to their habitat requirement. These assertions might complement based on results focused on the occupancy patterns vs. abundance dynamics and potential competition, which provide different views on the relationship between the two species. However, the results of occupancy are likely to be quite different if the study was covering a larger area into the distribution of both species.
Despite their ecological similarities, the coexistence between leopard cats and red foxes is made possible by a number of variables. These include seasonal resource availability, human interferences (Rota et al., 2016), and changes in seasonal patterns (Barrull et al., 2014). Coexisting carnivore populations can occupy various habitats by alternating between urban centers and core wilderness areas in most conserved areas with human activity (Vitekere et al., 2020). When dominant and subordinate species coexist in a community, the former may exclude the subordinate species from other locations (Noss et al., 1996; Creel, 2001). According to studies of a diverse set of ecologically important carnivore species
Table 5: The red fox, leopard cat, and yellow-throated marten pairwise conditional occupancy estimates within the study area are presented in the table. Each row represents a combination of species, and the conditional occupancy estimates, standard error, and significance for each species relative to the others are reported in each column.

Table 6: The observed Pianka’s index values for the niche overlap between the yellow-throated marten, red fox, and leopard cat in this study area are presented in the table. Pianka’s index determined the degree of spatial niche overlap between two species, with values ranging from 0 (showing no overlap) to 1 (representing complete overlap).
(Palomares & Caro, 1999), the distribution and abundance of carnivores can be significantly impacted by interspecific competition (Creel, 2001), and predicting the consequences of competition on carnivore populations can sometimes be challenging.
It is evident from various studies that interference may have an inverse density-dependent impact on the population’s growth, which would also reduce population increase at low densities. There is evidence from “natural experiments” that changes in one carnivore species’ density can significantly impact the density of other species through interference elimination (Henke & Bryant, 1999). One species will only be able to persist at a decreased density if it is forced to avoid or minimize its utilization of habitat patches due to the presence of another carnivore species (Linnell & Strand, 2000). The main distinction between interference and exploitative competition might be that interference reduces population growth and boosts variability at low densities. The SECR results indicate that red foxes may have a competitive advantage over leopard cats due to their higher estimated density. Evidence suggests that through interference competition, shifts in the population of one carnivore species, like red foxes, can significantly impact the density of other carnivore species, like leopard cats. Thus, leopard cats might persist at a lower density due to habitat avoidance or restricted habitat use when the red fox population is at a comparatively higher density. Results also provide insights into how dominating small carnivores can cause spatial turnover in other small carnivore guild members.
Carnivores coexist in some ecosystems, but interfering carnivores can reduce the impact of interspecific competition by segregating themselves temporally (Johnson et al., 1996). An adjunct method to studies on the abundance and density of species is estimating species trends with occupancy models. The use of camera traps assesses species’ status and tracks variations in their distribution range. Our study demonstrates the significance of a two-step occupancy modeling structure for generating initial, broad insights into the dynamics of small carnivore communities. We stress that the processes underpinning proposed direct and indirect ways should be tested through additional fine-scale investigations because interpreting insufficient estimates as observed abundances can result in overestimations (Jachowski & Harris, 2023). Our occupancy results suggest a substantial overlap between red fox and leopard cat. However, to investigate possible impacts on the abundances, densities, and occupancy status of various species, we initially included environmental covariates in the modeling process for SECR, N-Mixture, and occupancy models. These covariates were not included in the chosen best models, which were null models for N-Mixture and intercept-only models for occupancy. The smaller area and fewer captures might have impacted the selection, which may have impacted model convergence and the outcomes that followed.
In the context of resource partitioning and competition within their shared habitats, the results of our study provide insight into the potential repercussions of leopard cat (Prionailurus bengalensis) behaviors towards yellow-throated martens (Martes flavigula) and red foxes (Vulpes vulpes). The presence of red foxes in the ecosystem may be dominant, as indicated by their higher density and abundance than the other two species. This could have consequences for resource competition. The possibility of interspecific interactions is further highlighted by the observed overlap in space and time between red foxes and leopard cats, which suggests possible competition for resources like prey and habitat. As leopard cat and red fox are nocturnal, it is apparent that they show significant temporal overlaps (D4= 0.765). Our initial hypothesis is validated since our results strongly support the notion that leopard cats avoided foxes by habitat segregation, and the same is also evident from abundance and density estimates.
We acknowledge the possibility that the larger grid size utilized in the study may have contributed to lower detection chances. While this factor was not addressed, it could have influenced the results by affecting the detection probability of species within each grid cell. Further investigation into the potential impact of grid size on detection probabilities could provide valuable insights into the study’s findings. These findings have two significant implications for the conservation of small carnivores and the management of their habitats. Different habitat preferences of species suggest that maintaining habitat heterogeneity is essential for the conservation of different carnivore assemblages. Indicators of the integrity of the forest can be found in the varying ratios of small carnivores through time and space, which may reveal the degree of habitat degradation in the forest based on their varying affinities for mature forests.
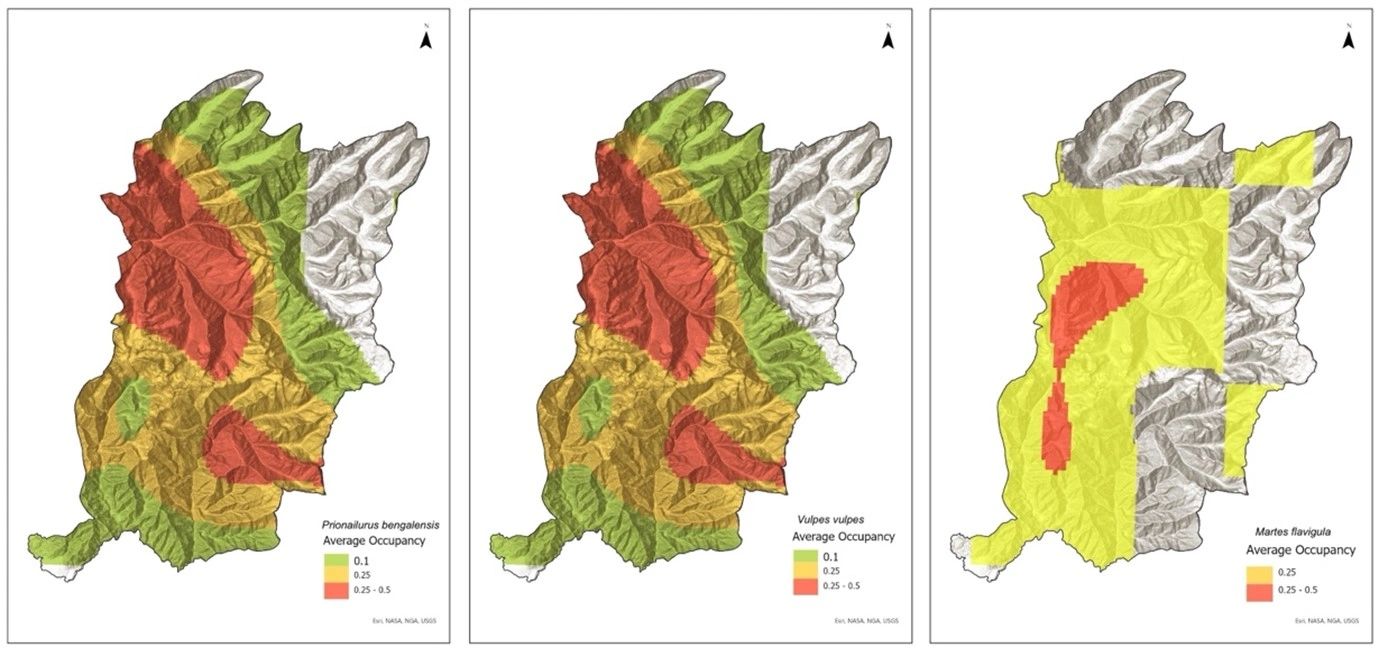
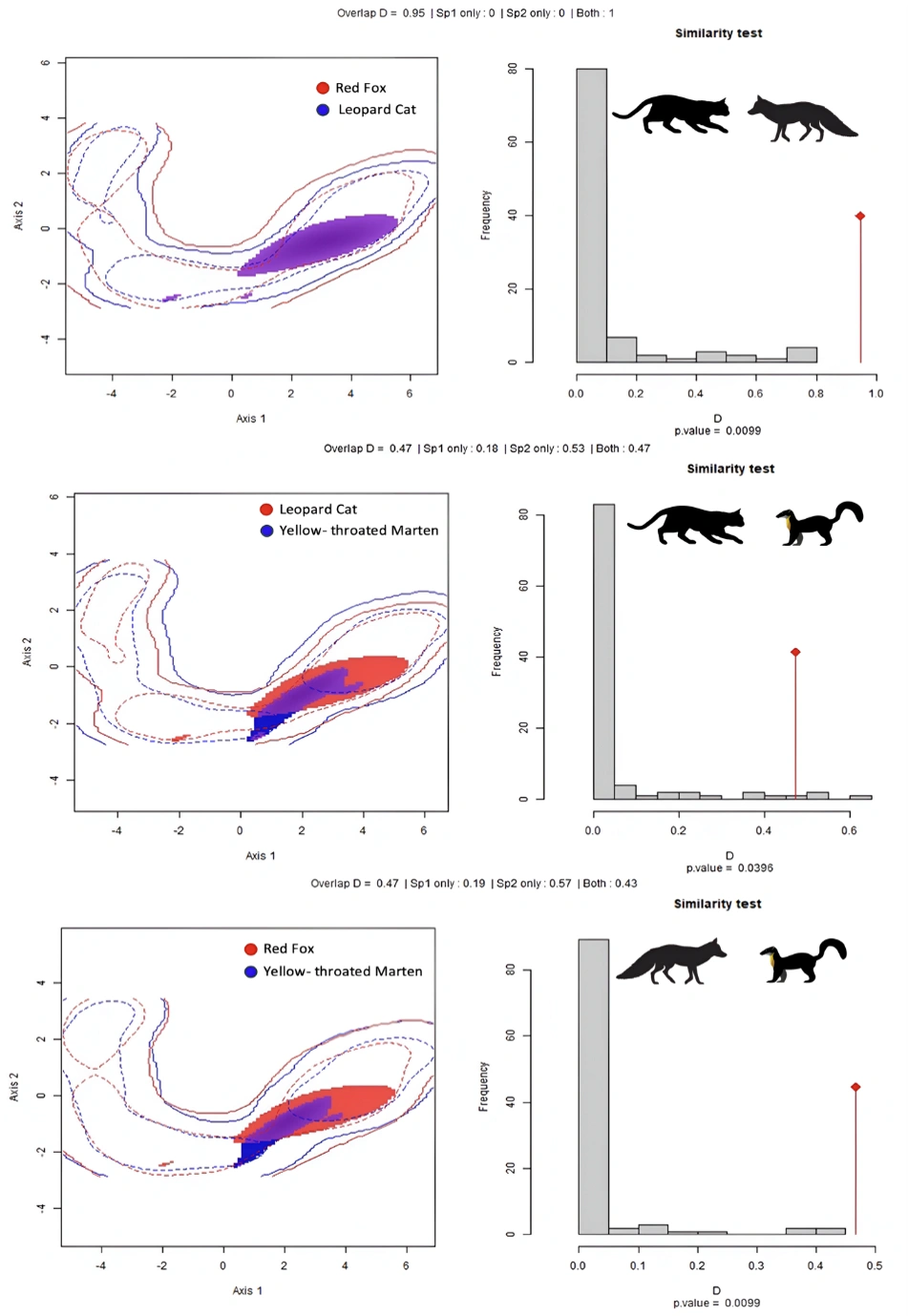
Figure 6: The average occupancy of red fox, yellow-throated marten, and leopard cat in the study area, along with pairwise overlaps in occupancy. There are similarities in the spatial distribution of leopard cat and red fox.
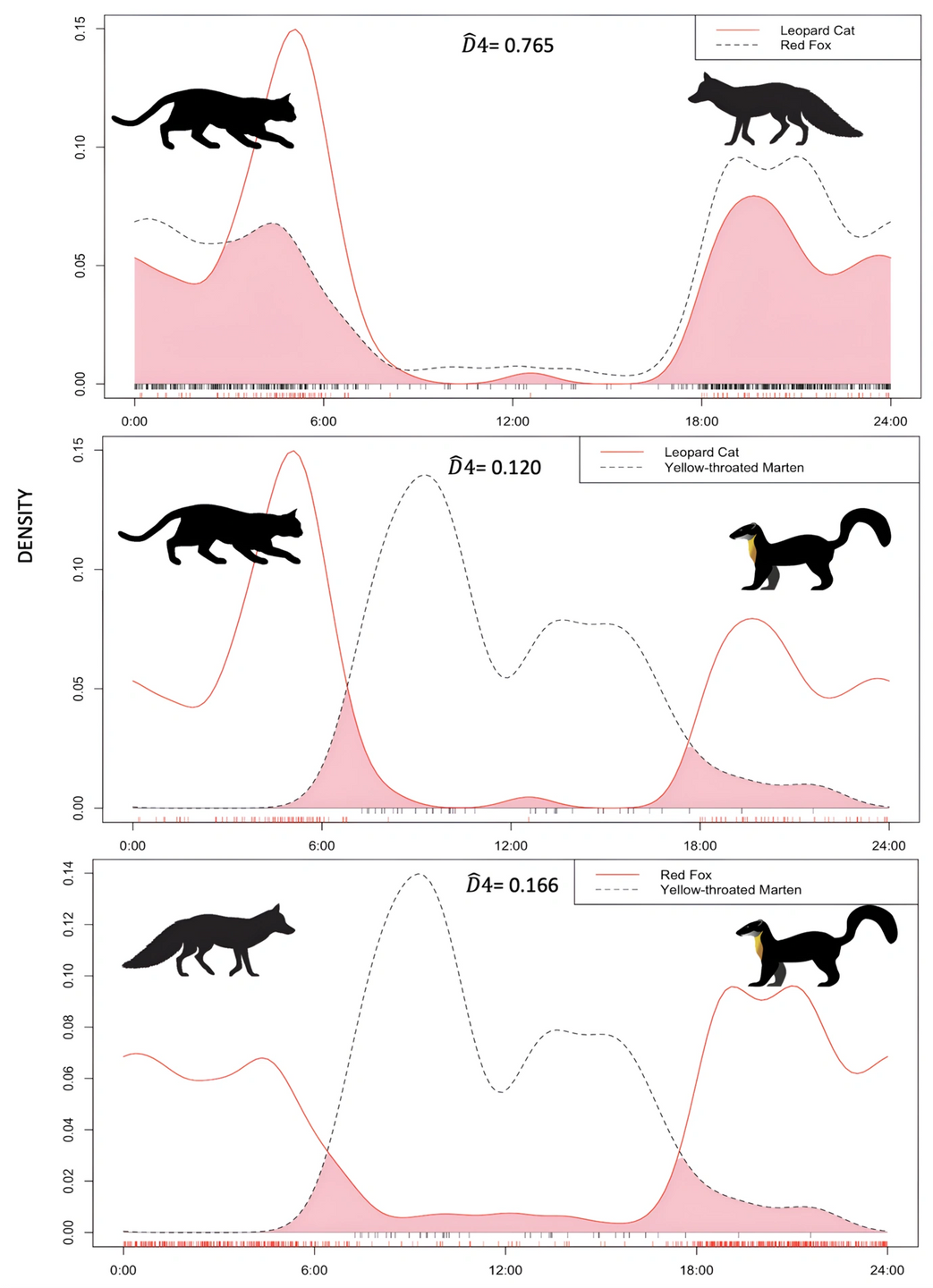
Figure 7: Pairwise comparisons between daily activity patterns of leopard cats, red foxes and yellow- throated martens. On the top left of each panel, D4 activity indices were obtained by comparing the patterns, with 95% confidence intervals
Acknowledgement
This study was funded by the Japan International Co-operation Agency (JICA) assisted project IHPFEM&L. The authors are grateful to the Director and Dean, for their encouragement and support. Special thanks are due to Himachal Pradesh Forest Department for their permission and support during fieldwork.
TO DOWNLOAD SUPPLEMENTARY MATERIAL CLICK HERE
CONFLICT OF INTEREST
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
DATA AVAILABILITY
The original contributions generated for the study are included in the article, further inquiries can be directed to the corresponding author/s.
AUTHOR CONTRIBUTIONS
TP- Conceptualization, Methodology, Data Curation, Formal Analysis, Investigation, Software, Writing-original draft, Writing- review and editing.
SL- Conceptualization, Methodology, Funding Acquisition, Project Administration, Supervision, Validation.
Edited By
Bilal Habib
Wildlife Institute of India
*CORRESPONDENCE
Salvador Lyngdoh
✉ salvador@wii.gov.in
CITATION
Parab, T., Lyngdoh, S. (2024) Understanding intra-guild dynamics: Do the presence of red foxes and yellow-throated martens shape the ecology of leopard cats in the lesser Himalayas? Journal of Wildlife Science,1 (1), 40-5.
https://doi.org/10.63033/JWLS.SBGZ2233
FUNDING
This study was funded by the Japan International Co-operation Agency (JICA) assisted project IHPFEM&L.
COPYRIGHT
© 2024 Parab, Lyngdoh. This is an open-access article, immediately and freely available to read, download, and share. The information contained in this article is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0![]()
![]() ), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepted academic practice. Copyright is retained by the author(s).
), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepted academic practice. Copyright is retained by the author(s).
PUBLISHED BY
Wildlife Institute of India, Dehradun, 248 001 INDIA
PUBLISHER'S NOTE
The Publisher, Journal of Wildlife Science or Editors cannot be held responsible for any errors or consequences arising from the use of the information contained in this article. All claims expressed in this article are solely those of the author(s) and do not necessarily represent those of their affiliated organisations or those of the publisher, the editors and the reviewers. Any product that may be evaluated or used in this article or claim made by its manufacturer is not guaranteed or endorsed by the publisher.
Akins, J. R. (2017). Distribution, genetic structure, and conservation status of the Cascade red fox in southern Washington. University of California, Davis.
Barrull, J., Mate, I., Ruiz-Olmo, J., Casanovas, J. G., Gosàlbez, J. & Salicrú, M. (2014). Factors and mechanisms that explain coexistence in a Mediterranean carnivore assemblage: an integrated study based on camera trapping and diet. Mammalian Biology, 79, 123-131.
https://doi.org/10.1016/j.mambio.2013.11.004
Bhatt, U., Singh Adhikari, B., Habib, B. & Lyngdoh, S. (2021). Temporal interactions and moon illumination effect on mammals in a tropical semievergreen forest of Manas National Park, Assam, India. Biotropica, 53(3), 831-845.
https://doi.org/10.1111/btp.12917
Bidlack, A. L., Merenlender, A., & Getz, W. M. (2008). Distribution of nonnative red foxes in East Bay oak woodlands. US For Serv Gen Tech Rep PSW, 217, 541-548.
Bischof, R., Ali, H., Kabir, M., Hameed, S. & Nawaz, M. A. (2014). Being the underdog: an elusive small carnivore uses space with prey and time without enemies. Journal of Zoology, 293(1), 40-48.
https://doi.org/10.1111/jzo.12100
Borchers, D. L., & Efford, M. G. (2008). Spatially explicit maximum likelihood methods for capture–recapture studies. Biometrics, 64(2), 377-385.
https://doi.org/10.1111/j.1541-0420.2007.00927.x
Bothma, J. D. P., Nel, J. A. J. & Macdonald, A. (1984). Food niche separation between four sympatric Namib Desert carnivores. Journal of Zoology, 202(3), 327-340.
https://doi.org/10.1111/j.1469-7998.1984.tb05086.x
Chiang, P. J., Pei, K. J. C., Vaughan, M. R. & Li, C. F. (2012). Niche relationships of carnivores in a subtropical primary forest in southern Taiwan. Zoological Studies, 51(4), 500-511.
Choi, M. B., Woo, D. & Choi, T. Y. (2015). Composition of the insect diet in feces of yellow-throated marten, Martes flavigula, in Jirisan National Park, South Korea. Journal of Ecology and Environment, 38(3), 389-395.
https://doi.org/10.5141/ecoenv.2015.041
Creel, S. (2001). Four factors modifying the effect of competition on carnivore population dynamics as illustrated by African wild dogs. Conservation Biology, 15(1), 271-274.
https://doi.org/10.1111/j.1523-1739.2001.99534.x
Creel, S. (2001). Interspecific competition and the population biology of extinction-prone carnivores. Carnivore conservation, 35-60.
Cristescu, B., Bernard, R. T. & Krause, J. (2013). Partitioning of space, habitat, and timing of activity by large felids in an enclosed South African system. Journal of Ethology, 31, 285-298.
https://doi.org/10.1007/s10164-013-0376-y
Di Bitetti, M. S., De Angelo, C. D., Di Blanco, Y. E. & Paviolo, A. (2010). Niche partitioning and species coexistence in a Neotropical felid assemblage. Acta Oecologica, 36(4), 403-412.
https://doi.org/10.1016/j.actao.2010.04.001
Dillon, A. & Kelly, M. J. (2008). Ocelot home range, overlap and density: comparing radio telemetry with camera trapping. Journal of Zoology, 275(4), 391-398.
https://doi.org/10.1111/j.1469-7998.2008.00452.x
Donadio, E. & Buskirk, S. W. (2006). Diet, morphology, and interspecific killing in Carnivora. The American Naturalist, 167(4), 524-536.
https://doi.org/10.1086/501033
Dunstone, N., Durbin, L., Wyllie, I., Freer, R., Jamett, G. A., Mazzolli, M. & Rose, S. (2002). Spatial organization, ranging behaviour and habitat use of the kodkod (Oncifelis guigna) in southern Chile. Journal of Zoology, 257(1), 1-11.
https://doi.org/10.1017/S0952836902000602
Efford, M. G. (2012). secr: Spatially explicit capture-reapture models. R package version 2.3. 2. https://CRAN. R-project. org/package= secr.
Elias, S. A. (2015). Differential insect and mammalian response to Late Quaternary climate change in the Rocky Mountain region of North America. Quaternary Science Reviews, 120, 57-70.
https://doi.org/10.1016/j.quascirev.2015.04.026
Fiske, I. & Chandler, R. (2011). Unmarked: an R package for fitting hierarchical models of wildlife occurrence and abundance. Journal of statistical software, 43, 1-23.
https://doi.org/10.18637/jss.v043.i10
Gilbert, N. A., Clare, J. D., Stenglein, J. L. & Zuckerberg, B. (2021). Abundance estimation of unmarked animals based on camera‐trap data. Conservation Biology, 35(1), 88-100.
https://doi.org/10.1111/cobi.13517
Grassel, S. M., Rachlow, J. L. & Williams, C. J. (2015). Spatial interactions between sympatric carnivores: asymmetric avoidance of an intraguild predator. Ecology and Evolution, 5(14), 2762-2773.
https://doi.org/10.1002/ece3.1561
Grassman, L. I., Tewes, M. E., Silvy, N. J. & Kreetiyutanont, K. (2005). Ecology of three sympatric felids in a mixed evergreen forest in north-central Thailand. Journal of mammalogy, 86(1), 29-38.
https://doi.org/10.1644/1545-1542(2005)086<0029:EOTSFI>2.0.CO;2
Harrison, D. J., Fuller, A. K. & Proulx, G. (Eds.). (2004). Martens and fishers (Martes) in human-altered environments: An international perspective. Springer Science & Business Media.
https://doi.org/10.1007/b99487
Hendry, A., Amir, Z., Decoeur, H., Mendes, C. P., Moore, J. H., Sovie, A. & Luskin, M. S. (2023). Marbled cats in Southeast Asia: are diurnal and semi‐arboreal felids at greater risk from human disturbances? Ecosphere, 14(1), e4338.
https://doi.org/10.1002/ecs2.4338
Henke, S. E. & Bryant, F. C. (1999). Effects of coyote removal on the faunal community in western Texas.The Journal of Wildlife Management, 1066-1081.
https://doi.org/10.2307/3802826
Howe, E. J., Potter, D., Beauclerc, K. B., Jackson, K. E. & Northrup, J. M. (2022). Estimating animal abundance at multiple scales by spatially explicit capture–recapture. Ecological Applications, 32(7), e2638.
https://doi.org/10.1002/eap.2638
Hua, Y., Vitekere, K., Wang, J., Zhu, M., Zaman, M. & Jiang, G. (2020, May). Coexistence of sympatric carnivores in a relatively homogenous landscape and the effects of environmental factors on site occupation. In Annales Zoologici Fennici (Vol. 57, No. 1-6, 47-58). Finnish Zoological and Botanical Publishing Board.
https://doi.org/10.5735/086.057.0106
Jachowski, D. S., Marneweck, C. J., Olfenbuttel, C., & Harris, S. N. (2024). Support for the size‐mediated sensitivity hypothesis within a diverse carnivore community. Journal of Animal Ecology, 93(1), 109-122.
https://doi.org/10.1111/1365-2656.13916
Ji, C., Li, H. D., Xiao, W., Xu, K., Ren, Y., Li, H., ... & Xiao, Z. (2023). Fine-scale interactions between leopard cats and their potential prey with contrasting diel activities in a livestock-dominated nature reserve. Animals, 13(8), 1296.
https://doi.org/10.3390/ani13081296
Johnson, W. E., Fuller, T. K. & Franklin, W. L. (1996). Sympatry in canids: a review and assessment. Carnivore behavior, ecology, and evolution, 2, 189-218.
Karanth, K. U. & Chellam, R. (2009). Carnivore conservation at the crossroads. Oryx, 43(1), 1-2.
https://doi.org/10.1017/S003060530843106X
Kery, M., Gardner, B., Stoeckle, T., Weber, D. & Royle, J. A. (2011). Use of spatial capture‐recapture modeling and DNA data to estimate densities of elusive animals. Conservation Biology, 25(2), 356-364.
https://doi.org/10.1111/j.1523-1739.2010.01616.x
Lantschner, M. V., Rusch, V., & Hayes, J. P. (2012). Habitat use by carnivores at different spatial scales in a plantation forest landscape in Patagonia, Argentina. Forest Ecology and Management, 269, 271-278.
https://doi.org/10.1016/j.foreco.2011.12.045
Laundré, J. W., Hernández, L. & Ripple, W. J. (2010). The landscape of fear: ecological implications of being afraid. The open ecology journal, 3(1).
https://doi.org/10.2174/1874213001003030001
Linnell, J. D. & Strand, O. (2000). Interference interactions, coexistence and conservation of mammalian carnivores. Diversity and Distributions, 6(4), 169-176.
https://doi.org/10.1046/j.1472-4642.2000.00069.x
MacArthur, R. & Levins, R. (1967). The limiting similarity, convergence, and divergence of coexisting species. The American Naturalist, 101(921), 377-385.
https://doi.org/10.1086/282505
Macdonald, D. W. (1983). The ecology of carnivore social behaviour. Nature, 301(5899), 379-384.
https://doi.org/10.1038/301379a0
Macdonald D.W., Reynolds J.C. (2008) Vulpes vulpes. IUCN 2011. IUCN Red list of threatened species. Version 2011.2. HYPERLINK B https://www.iucnredlist.org ; www.iucnredlist.org.
MacKenzie, D. I., Nichols, J. D., Royle, J. A., Pollock, K. H., Bailey, L. & Hines, J. E. (2017). Occupancy estimation and modeling: inferring patterns and dynamics of species occurrence. Elsevier
https://doi.org/10.1016/B978-0-12-407197-1.00019-3
Marcella, J. K. (2004). Sunquist, M., F. Sunquist. 2002. Wild Cats of the World. University of Chicago Press, Chicago and London. 452 pp. ISBN 0-226-77999-8 (hardbound) $45.00.
McHugh, D., Goldingay, R. L. & Letnic, M. (2022). Occupancy and co-occurrence patterns of endemic mammals and introduced predators across a broad geographical gradient in eastern Australia. Biodiversity and Conservation, 31(3), 989-1021.
https://doi.org/10.1007/s10531-022-02374-0
Meredith, M. & Ridout, M. S. (2014). The overlap package.
Mohamed, A., Sollmann, R., Bernard, H., Ambu, L. N., Lagan, P., Mannan, S., ... & Wilting, A. (2013). Density and habitat use of the leopard cat (Prionailurus bengalensis) in three commercial forest reserves in Sabah, Malaysian Borneo. Journal of mammalogy, 94(1), 82-89.
https://doi.org/10.1644/11-MAMM-A-394.1
Mohd-Azlan, J., Kaicheen, S. S., Hong, L. L. C., Yi, M. C. K., Maiwald, M. J., Helmy, O. E., ... & Brodie, J. F. (2023). Ecology, occurrence and distribution of wild felids in Sarawak, Malaysian Borneo. Oryx, 57(2), 252-261.
https://doi.org/10.1017/S0030605321001484
Mowry, S., Pendleton, J., Keesing, F., Teator, M. & Ostfeld, R. S. (2022). Estimates of wildlife species richness, occupancy, and habitat preference in a human-impacted landscape in New York State.
https://doi.org/10.21203/rs.3.rs-647329/v1
Nandini, R. & Karthik, T. (2007). Field observations of Yellow- throated Martens Martes flavigula feeding on flowers in Meghalaya, North-east India. Small Carnivore Conservation, 37, 26-27.
Noss, R. F., Quigley, H. B., Hornocker, M. G., Merrill, T. & Paquet, P. C. (1996). Conservation Biology and carnivore conservation in the Rocky Mountains. Conservation Biology, 10(4), 949-963.
https://doi.org/10.1046/j.1523-1739.1996.10040949.x
Nowak, R. M. (1999). Walker's Mammals of the World (Vol. 1). JHU press.
https://doi.org/10.56021/9780801857898
Nowell, K. & Jackson, P. (Eds.). (1996). Wild cats: status survey and conservation action plan (Vol. 382). Gland, Switzerland: IUCN.
Palomares, F. & Caro, T. M. (1999). Interspecific killing among mammalian carnivores. The American Naturalist, 153(5), 492-508.
https://doi.org/10.1086/303189
Patel, R. P., Wutke, S., Lenz, D., Mukherjee, S., Ramakrishnan, U., Veron, G., ... & Förster, D. W. (2017). Genetic structure and phylogeography of the leopard cat (Prionailurus bengalensis) inferred from mitochondrial genomes. Journal of Heredity, 108(4), 349-360.
https://doi.org/10.1093/jhered/esx017
Pozzanghera, C. (2015). Non-Invasive methods for obtaining occupancy probabilities and density estimates of interior Alaska's mesocarnivore populations. University of Alaska Fairbanks.
R Core Team. (2016). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org
Reppucci, J., Gardner, B. & Lucherini, M. (2011). Estimating detection and density of the Andean cat in the high Andes. Journal of mammalogy, 92(1), 140-147.
https://doi.org/10.1644/10-MAMM-A-053.1
Rodgers, W. A., & Panwar, H. S. (1988). Planning a wildlife protected area network in India.
Rota, C. T., Ferreira, M. A., Kays, R. W., Forrester, T. D., Kalies, E. L., McShea, W. J., ... & Millspaugh, J. J. (2016). A multispecies occupancy model for two or more interacting species. Methods in Ecology and Evolution, 7(10), 1164-1173.
https://doi.org/10.1111/2041-210X.12587
Rota, C. T., Wikle, C. K., Kays, R. W., Forrester, T. D., McShea, W. J., Parsons, A. W. & Millspaugh, J. J. (2016). A two‐species occupancy model accommodating simultaneous spatial and interspecific dependence. Ecology, 97(1), 48-53.
https://doi.org/10.1890/15-1193.1
Salvatori, M., Tenan, S., Oberosler, V., Augugliaro, C., Christe, P., Groff, C., ... & Rovero, F. (2021). Co-occurrence of snow leopard, wolf and Siberian ibex under livestock encroachment into protected areas across the Mongolian Altai. Biological Conservation, 261, 109294.
https://doi.org/10.1016/j.biocon.2021.109294
Sarmento, P., Cruz, J., Eira, C. & Fonseca, C. (2009). Evaluation of camera trapping for estimating red fox abundance. The Journal of Wildlife Management, 73(7), 1207-1212.
https://doi.org/10.2193/2008-288
Seidensticker, J. (1976). On the ecological separation between tigers and leopards. Biotropica, 225-234.
https://doi.org/10.2307/2989714
Silmi, M., Putra, K., Amran, A., Huda, M., Fanani, A. F., Galdikas, B. M., ... & Traeholt, C. (2021). Activity and ranging behavior of leopard cats (Prionailurus bengalensis) in an oil palm landscape. Frontiers in Environmental Science, 9, 651939.
https://doi.org/10.3389/fenvs.2021.651939
Skinner, E. B., Glidden, C. K., MacDonald, A. J., & Mordecai, E. A. (2023). Human footprint is associated with shifts in the assemblages of major vector-borne diseases. Nature sustainability, 6(6), 652-661.
https://doi.org/10.1038/s41893-023-01080-1
Sliwa, A. (2004). Home range size and social organisation of black-footed cats. Mammalian Biology, 69(2), 96-107.
https://doi.org/10.1078/1616-5047-00124
Stantial, M. L., Cohen, J. B., Darrah, A. J., Farrell, S. & Maslo, B. (2020). Red fox use of landscapes with nesting shorebirds. The Journal of Wildlife Management, 84(8), 1536-1547.
https://doi.org/10.1002/jwmg.21949
Sunquist, M. E. & Sunquist, F. C. (1989). Ecological constraints on predation by large felids. In Carnivore behavior, ecology, and evolution (283-301). Boston, MA: Springer US.
https://doi.org/10.1007/978-1-4757-4716-4_11
Taylor, S. L. & Buskirk, S. W. (1994). Forest microenvironments and resting energetics of the American marten Martes americana. Ecography, 17(3), 249-256.
https://doi.org/10.1111/j.1600-0587.1994.tb00100.x
Trolle, M. & Kéry, M. (2003). Estimation of ocelot density in the Pantanal using capture-recapture analysis of camera-trapping data. Journal of mammalogy, 84(2), 607-614.
https://doi.org/10.1644/1545-1542(2003)084<0607:EOODIT>2.0.CO;2
Vitekere, K., Wang, J., Karanja, H., Consolée, K. T., Jiang, G. & Hua, Y. (2020). Dynamic in species estimates of carnivores (leopard cat, red fox, and north Chinese leopard): A multi-year assessment of occupancy and coexistence in the Tieqiaoshan Nature Reserve, Shanxi Province, China. Animals, 10(8), 1333.
https://doi.org/10.3390/ani10081333
Walton, Z., Samelius, G., Odden, M. & Willebrand, T. (2017). Variation in home range size of red foxes Vulpes vulpes along a gradient of productivity and human landscape alteration. PloS one, 12(4), e0175291.
https://doi.org/10.1371/journal.pone.0175291
Zhang, J., Ding, Q. & Huang, J. (2016). span: Species association analysis. R package version 0.2, 2, 33.

Edited By
Bilal Habib
Wildlife Institute of India
*CORRESPONDENCE
Salvador Lyngdoh
✉ salvador@wii.gov.in
CITATION
Parab, T., Lyngdoh, S. (2024) Understanding intra-guild dynamics: Do the presence of red foxes and yellow-throated martens shape the ecology of leopard cats in the lesser Himalayas? Journal of Wildlife Science,1 (1), 40-5.
https://doi.org/10.63033/JWLS.SBGZ2233
FUNDING
This study was funded by the Japan International Co-operation Agency (JICA) assisted project IHPFEM&L.
COPYRIGHT
© 2024 Parab, Lyngdoh. This is an open-access article, immediately and freely available to read, download, and share. The information contained in this article is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0![]()
![]() ), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepted academic practice. Copyright is retained by the author(s).
), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepted academic practice. Copyright is retained by the author(s).
PUBLISHED BY
Wildlife Institute of India, Dehradun, 248 001 INDIA
PUBLISHER'S NOTE
The Publisher, Journal of Wildlife Science or Editors cannot be held responsible for any errors or consequences arising from the use of the information contained in this article. All claims expressed in this article are solely those of the author(s) and do not necessarily represent those of their affiliated organisations or those of the publisher, the editors and the reviewers. Any product that may be evaluated or used in this article or claim made by its manufacturer is not guaranteed or endorsed by the publisher.
Akins, J. R. (2017). Distribution, genetic structure, and conservation status of the Cascade red fox in southern Washington. University of California, Davis.
Barrull, J., Mate, I., Ruiz-Olmo, J., Casanovas, J. G., Gosàlbez, J. & Salicrú, M. (2014). Factors and mechanisms that explain coexistence in a Mediterranean carnivore assemblage: an integrated study based on camera trapping and diet. Mammalian Biology, 79, 123-131.
https://doi.org/10.1016/j.mambio.2013.11.004
Bhatt, U., Singh Adhikari, B., Habib, B. & Lyngdoh, S. (2021). Temporal interactions and moon illumination effect on mammals in a tropical semievergreen forest of Manas National Park, Assam, India. Biotropica, 53(3), 831-845.
https://doi.org/10.1111/btp.12917
Bidlack, A. L., Merenlender, A. & Getz, W. M. (2008). Distribution of nonnative red foxes in East Bay oak woodlands. US For Serv Gen Tech Rep PSW, 217, 541-548.
Bischof, R., Ali, H., Kabir, M., Hameed, S. & Nawaz, M. A. (2014). Being the underdog: an elusive small carnivore uses space with prey and time without enemies. Journal of Zoology, 293(1), 40-48.
https://doi.org/10.1111/jzo.12100
Borchers, D. L., & Efford, M. G. (2008). Spatially explicit maximum likelihood methods for capture–recapture studies. Biometrics, 64(2), 377-385.
https://doi.org/10.1111/j.1541-0420.2007.00927.x
Bothma, J. D. P., Nel, J. A. J. & Macdonald, A. (1984). Food niche separation between four sympatric Namib Desert carnivores. Journal of Zoology, 202(3), 327-340.
https://doi.org/10.1111/j.1469-7998.1984.tb05086.x
Chiang, P. J., Pei, K. J. C., Vaughan, M. R. & Li, C. F. (2012). Niche relationships of carnivores in a subtropical primary forest in southern Taiwan. Zoological Studies, 51(4), 500-511.
Choi, M. B., Woo, D. & Choi, T. Y. (2015). Composition of the insect diet in feces of yellow-throated marten, Martes flavigula, in Jirisan National Park, South Korea. Journal of Ecology and Environment, 38(3), 389-395.
https://doi.org/10.5141/ecoenv.2015.041
Creel, S. (2001). Four factors modifying the effect of competition on carnivore population dynamics as illustrated by African wild dogs. Conservation Biology, 15(1), 271-274.
https://doi.org/10.1111/j.1523-1739.2001.99534.x
Creel, S. (2001). Interspecific competition and the population biology of extinction-prone carnivores. Carnivore conservation, 35-60.
Cristescu, B., Bernard, R. T. & Krause, J. (2013). Partitioning of space, habitat, and timing of activity by large felids in an enclosed South African system. Journal of Ethology, 31, 285-298.
https://doi.org/10.1007/s10164-013-0376-y
Di Bitetti, M. S., De Angelo, C. D., Di Blanco, Y. E. & Paviolo, A. (2010). Niche partitioning and species coexistence in a Neotropical felid assemblage. Acta Oecologica, 36(4), 403-412.
https://doi.org/10.1016/j.actao.2010.04.001
Dillon, A. & Kelly, M. J. (2008). Ocelot home range, overlap and density: comparing radio telemetry with camera trapping. Journal of Zoology, 275(4), 391-398.
https://doi.org/10.1111/j.1469-7998.2008.00452.x
Donadio, E. & Buskirk, S. W. (2006). Diet, morphology, and interspecific killing in Carnivora. The American Naturalist, 167(4), 524-536.
https://doi.org/10.1086/501033
Dunstone, N., Durbin, L., Wyllie, I., Freer, R., Jamett, G. A., Mazzolli, M. & Rose, S. (2002). Spatial organization, ranging behaviour and habitat use of the kodkod (Oncifelis guigna) in southern Chile. Journal of Zoology, 257(1), 1-11.
https://doi.org/10.1017/S0952836902000602
Efford, M. G. (2012). secr: Spatially explicit capture-reapture models. R package version 2.3. 2. https://CRAN. R-project. org/package= secr.
Elias, S. A. (2015). Differential insect and mammalian response to Late Quaternary climate change in the Rocky Mountain region of North America. Quaternary Science Reviews, 120, 57-70.
https://doi.org/10.1016/j.quascirev.2015.04.026
Fiske, I. & Chandler, R. (2011). Unmarked: an R package for fitting hierarchical models of wildlife occurrence and abundance. Journal of statistical software, 43, 1-23.
https://doi.org/10.18637/jss.v043.i10
Gilbert, N. A., Clare, J. D., Stenglein, J. L. & Zuckerberg, B. (2021). Abundance estimation of unmarked animals based on camera‐trap data. Conservation Biology, 35(1), 88-100.
https://doi.org/10.1111/cobi.13517
Grassel, S. M., Rachlow, J. L. & Williams, C. J. (2015). Spatial interactions between sympatric carnivores: asymmetric avoidance of an intraguild predator. Ecology and Evolution, 5(14), 2762-2773.
https://doi.org/10.1002/ece3.1561
Grassman, L. I., Tewes, M. E., Silvy, N. J. & Kreetiyutanont, K. (2005). Ecology of three sympatric felids in a mixed evergreen forest in north-central Thailand. Journal of mammalogy, 86(1), 29-38.
https://doi.org/10.1644/1545-1542(2005)086<0029:EOTSFI>2.0.CO;2
Harrison, D. J., Fuller, A. K. & Proulx, G. (Eds.). (2004). Martens and fishers (Martes) in human-altered environments: An international perspective. Springer Science & Business Media.
https://doi.org/10.1007/b99487
Hendry, A., Amir, Z., Decoeur, H., Mendes, C. P., Moore, J. H., Sovie, A. & Luskin, M. S. (2023). Marbled cats in Southeast Asia: are diurnal and semi‐arboreal felids at greater risk from human disturbances? Ecosphere, 14(1), e4338.
https://doi.org/10.1002/ecs2.4338
Henke, S. E. & Bryant, F. C. (1999). Effects of coyote removal on the faunal community in western Texas.The Journal of Wildlife Management, 1066-1081.
https://doi.org/10.2307/3802826
Howe, E. J., Potter, D., Beauclerc, K. B., Jackson, K. E. & Northrup, J. M. (2022). Estimating animal abundance at multiple scales by spatially explicit capture–recapture. Ecological Applications, 32(7), e2638.
https://doi.org/10.1002/eap.2638
Hua, Y., Vitekere, K., Wang, J., Zhu, M., Zaman, M. & Jiang, G. (2020, May). Coexistence of sympatric carnivores in a relatively homogenous landscape and the effects of environmental factors on site occupation. In Annales Zoologici Fennici (Vol. 57, No. 1-6, 47-58). Finnish Zoological and Botanical Publishing Board.
https://doi.org/10.5735/086.057.0106
Jachowski, D. S., Marneweck, C. J., Olfenbuttel, C., & Harris, S. N. (2024). Support for the size‐mediated sensitivity hypothesis within a diverse carnivore community. Journal of Animal Ecology, 93(1), 109-122.
https://doi.org/10.1111/1365-2656.13916
Ji, C., Li, H. D., Xiao, W., Xu, K., Ren, Y., Li, H., ... & Xiao, Z. (2023). Fine-scale interactions between leopard cats and their potential prey with contrasting diel activities in a livestock-dominated nature reserve. Animals, 13(8), 1296.
https://doi.org/10.3390/ani13081296
Johnson, W. E., Fuller, T. K. & Franklin, W. L. (1996). Sympatry in canids: a review and assessment. Carnivore behavior, ecology, and evolution, 2, 189-218.
Karanth, K. U. & Chellam, R. (2009). Carnivore conservation at the crossroads. Oryx, 43(1), 1-2.
https://doi.org/10.1017/S003060530843106X
Kery, M., Gardner, B., Stoeckle, T., Weber, D. & Royle, J. A. (2011). Use of spatial capture‐recapture modeling and DNA data to estimate densities of elusive animals. Conservation Biology, 25(2), 356-364.
https://doi.org/10.1111/j.1523-1739.2010.01616.x
Lantschner, M. V., Rusch, V., & Hayes, J. P. (2012). Habitat use by carnivores at different spatial scales in a plantation forest landscape in Patagonia, Argentina. Forest Ecology and Management, 269, 271-278.
https://doi.org/10.1016/j.foreco.2011.12.045
Laundré, J. W., Hernández, L. & Ripple, W. J. (2010). The landscape of fear: ecological implications of being afraid. The open ecology journal, 3(1).
https://doi.org/10.2174/1874213001003030001
Linnell, J. D. & Strand, O. (2000). Interference interactions, coexistence and conservation of mammalian carnivores. Diversity and Distributions, 6(4), 169-176.
https://doi.org/10.1046/j.1472-4642.2000.00069.x
MacArthur, R. & Levins, R. (1967). The limiting similarity, convergence, and divergence of coexisting species. The American Naturalist, 101(921), 377-385.
https://doi.org/10.1086/282505
Macdonald, D. W. (1983). The ecology of carnivore social behaviour. Nature, 301(5899), 379-384.
https://doi.org/10.1038/301379a0
Macdonald D.W., Reynolds J.C. (2008) Vulpes vulpes. IUCN 2011. IUCN Red list of threatened species. Version 2011.2. HYPERLINK B https://www.iucnredlist.org ; www.iucnredlist.org.
MacKenzie, D. I., Nichols, J. D., Royle, J. A., Pollock, K. H., Bailey, L. & Hines, J. E. (2017). Occupancy estimation and modeling: inferring patterns and dynamics of species occurrence. Elsevier
https://doi.org/10.1016/B978-0-12-407197-1.00019-3
Marcella, J. K. (2004). Sunquist, M., F. Sunquist. 2002. Wild Cats of the World. University of Chicago Press, Chicago and London. 452 pp. ISBN 0-226-77999-8 (hardbound) $45.00.
McHugh, D., Goldingay, R. L. & Letnic, M. (2022). Occupancy and co-occurrence patterns of endemic mammals and introduced predators across a broad geographical gradient in eastern Australia. Biodiversity and Conservation, 31(3), 989-1021.
https://doi.org/10.1007/s10531-022-02374-0
Meredith, M. & Ridout, M. S. (2014). The overlap package.
Mohamed, A., Sollmann, R., Bernard, H., Ambu, L. N., Lagan, P., Mannan, S., ... & Wilting, A. (2013). Density and habitat use of the leopard cat (Prionailurus bengalensis) in three commercial forest reserves in Sabah, Malaysian Borneo. Journal of mammalogy, 94(1), 82-89.
https://doi.org/10.1644/11-MAMM-A-394.1
Mohd-Azlan, J., Kaicheen, S. S., Hong, L. L. C., Yi, M. C. K., Maiwald, M. J., Helmy, O. E., ... & Brodie, J. F. (2023). Ecology, occurrence and distribution of wild felids in Sarawak, Malaysian Borneo. Oryx, 57(2), 252-261.
https://doi.org/10.1017/S0030605321001484
Mowry, S., Pendleton, J., Keesing, F., Teator, M. & Ostfeld, R. S. (2022). Estimates of wildlife species richness, occupancy, and habitat preference in a human-impacted landscape in New York State.
https://doi.org/10.21203/rs.3.rs-647329/v1
Nandini, R. & Karthik, T. (2007). Field observations of Yellow- throated Martens Martes flavigula feeding on flowers in Meghalaya, North-east India. Small Carnivore Conservation, 37, 26-27.
Noss, R. F., Quigley, H. B., Hornocker, M. G., Merrill, T. & Paquet, P. C. (1996). Conservation Biology and carnivore conservation in the Rocky Mountains. Conservation Biology, 10(4), 949-963.
https://doi.org/10.1046/j.1523-1739.1996.10040949.x
Nowak, R. M. (1999). Walker's Mammals of the World (Vol. 1). JHU press.
https://doi.org/10.56021/9780801857898
Nowell, K. & Jackson, P. (Eds.). (1996). Wild cats: status survey and conservation action plan (Vol. 382). Gland, Switzerland: IUCN.
Palomares, F. & Caro, T. M. (1999). Interspecific killing among mammalian carnivores. The American Naturalist, 153(5), 492-508.
https://doi.org/10.1086/303189
Patel, R. P., Wutke, S., Lenz, D., Mukherjee, S., Ramakrishnan, U., Veron, G., ... & Förster, D. W. (2017). Genetic structure and phylogeography of the leopard cat (Prionailurus bengalensis) inferred from mitochondrial genomes. Journal of Heredity, 108(4), 349-360.
https://doi.org/10.1093/jhered/esx017
Pozzanghera, C. (2015). Non-Invasive methods for obtaining occupancy probabilities and density estimates of interior Alaska's mesocarnivore populations. University of Alaska Fairbanks.
R Core Team. (2016). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org
Reppucci, J., Gardner, B. & Lucherini, M. (2011). Estimating detection and density of the Andean cat in the high Andes. Journal of mammalogy, 92(1), 140-147.
https://doi.org/10.1644/10-MAMM-A-053.1
Rodgers, W. A., & Panwar, H. S. (1988). Planning a wildlife protected area network in India.
Rota, C. T., Ferreira, M. A., Kays, R. W., Forrester, T. D., Kalies, E. L., McShea, W. J., ... & Millspaugh, J. J. (2016). A multispecies occupancy model for two or more interacting species. Methods in Ecology and Evolution, 7(10), 1164-1173.
https://doi.org/10.1111/2041-210X.12587
Rota, C. T., Wikle, C. K., Kays, R. W., Forrester, T. D., McShea, W. J., Parsons, A. W. & Millspaugh, J. J. (2016). A two‐species occupancy model accommodating simultaneous spatial and interspecific dependence. Ecology, 97(1), 48-53.
https://doi.org/10.1890/15-1193.1
Salvatori, M., Tenan, S., Oberosler, V., Augugliaro, C., Christe, P., Groff, C., ... & Rovero, F. (2021). Co-occurrence of snow leopard, wolf and Siberian ibex under livestock encroachment into protected areas across the Mongolian Altai. Biological Conservation, 261, 109294.
https://doi.org/10.1016/j.biocon.2021.109294
Sarmento, P., Cruz, J., Eira, C. & Fonseca, C. (2009). Evaluation of camera trapping for estimating red fox abundance. The Journal of Wildlife Management, 73(7), 1207-1212.
https://doi.org/10.2193/2008-288
Seidensticker, J. (1976). On the ecological separation between tigers and leopards. Biotropica, 225-234.
https://doi.org/10.2307/2989714
Silmi, M., Putra, K., Amran, A., Huda, M., Fanani, A. F., Galdikas, B. M., ... & Traeholt, C. (2021). Activity and ranging behavior of leopard cats (Prionailurus bengalensis) in an oil palm landscape. Frontiers in Environmental Science, 9, 651939.
https://doi.org/10.3389/fenvs.2021.651939
Skinner, E. B., Glidden, C. K., MacDonald, A. J., & Mordecai, E. A. (2023). Human footprint is associated with shifts in the assemblages of major vector-borne diseases. Nature sustainability, 6(6), 652-661.
https://doi.org/10.1038/s41893-023-01080-1
Sliwa, A. (2004). Home range size and social organisation of black-footed cats. Mammalian Biology, 69(2), 96-107.
https://doi.org/10.1078/1616-5047-00124
Stantial, M. L., Cohen, J. B., Darrah, A. J., Farrell, S. & Maslo, B. (2020). Red fox use of landscapes with nesting shorebirds. The Journal of Wildlife Management, 84(8), 1536-1547.
https://doi.org/10.1002/jwmg.21949
Sunquist, M. E. & Sunquist, F. C. (1989). Ecological constraints on predation by large felids. In Carnivore behavior, ecology, and evolution (283-301). Boston, MA: Springer US.
https://doi.org/10.1007/978-1-4757-4716-4_11
Taylor, S. L. & Buskirk, S. W. (1994). Forest microenvironments and resting energetics of the American marten Martes americana. Ecography, 17(3), 249-256.
https://doi.org/10.1111/j.1600-0587.1994.tb00100.x
Trolle, M. & Kéry, M. (2003). Estimation of ocelot density in the Pantanal using capture-recapture analysis of camera-trapping data. Journal of mammalogy, 84(2), 607-614.
https://doi.org/10.1644/1545-1542(2003)084<0607:EOODIT>2.0.CO;2
Vitekere, K., Wang, J., Karanja, H., Consolée, K. T., Jiang, G. & Hua, Y. (2020). Dynamic in species estimates of carnivores (leopard cat, red fox, and north Chinese leopard): A multi-year assessment of occupancy and coexistence in the Tieqiaoshan Nature Reserve, Shanxi Province, China. Animals, 10(8), 1333.
https://doi.org/10.3390/ani10081333
Walton, Z., Samelius, G., Odden, M. & Willebrand, T. (2017). Variation in home range size of red foxes Vulpes vulpes along a gradient of productivity and human landscape alteration. PloS one, 12(4), e0175291.
https://doi.org/10.1371/journal.pone.0175291
Zhang, J., Ding, Q. & Huang, J. (2016). span: Species association analysis. R package version 0.2, 2, 33.