TYPE: Research Article ![]()
Urjit Bhatt¹ , Salvador Lyngdoh¹*
, Salvador Lyngdoh¹*
¹Department of Landscape level planning & Management, Wildlife Institute of India, Dehradun 248001, Uttarakhand, India

Introduction
Carnivores are integral components of mammal communities, and despite their naturally low densities, they often strongly influence other species in the community (Vernes et al. 2021). Their role is pivotal in shaping ecosystems, primarily through their top-down effects on prey species (Glen & Dickman 2005; Terborgh & Estes 2013; Ripple et al. 2014). Additionally, large carnivores can influence the dynamics of mesocarnivores by engaging in intraguild competition, thereby creating diverse food web pathways (Ripple et al. 2013; Suraci et al. 2016). Interspecific competition and resource partitioning have been identified as one of the most critical variables in structuring carnivore communities despite the wide range of factors that might affect carnivore populations (Di Bitetti et al. 2010; May et al. 2008). Thus, the occurrence of a particular species within a given environment results from a delicate trade-off between habitat quality and its ability to effectively compete with other species for resources, thereby directly or indirectly affecting resource access (Araujo & Guisan 2006; Soberón 2007). MacArthur & Levins (1967) suggested that two competitive species can co-exist if their niches differ in at least one or more dimensions. Such niche segregation occurs primarily along three axes of niche dimension: spatial (habitat), trophic, and temporal (Pianka 1974).
Ecologists studying carnivores in tropical rainforests struggle to observe elusive animals directly, often relying on indirect signs like tracks or scats, which provide limited insights (Sollmann et al. 2013). However, camera trapping has transformed carnivore research, offering more reliable evidence of their presence and facilitating the implementation of standardised sampling protocols (Sollmann et al. 2013). Tropical carnivores, with their body size (medium or large), morphology (natural marks for individual identification), guild/habitat in which they live (terrestrial—allowing for relatively simple placement of equipment), behaviour (readily use trails), secretiveness/ elusiveness (challenging to study with other methods), rarity (requiring considerable survey effort), activity patterns (often nocturnal and challenging for direct observation) and generally remote locations are well-suited for camera trapping surveys over traditional methods (Griffiths & Schaick 1993; Goldman & Winther-Hansen 2003; Wibisono & Mccarthy 2010; Bhatt et al. 2021, 2022, 2023a, 2023b).
A comprehensive understanding of the dynamics within tropical carnivore communities is of utmost importance for effectively managing and conserving these ecosystems (Rabinowitz & Walker 1991). The Manas National Park (MNP), situated in the Indo-Gangetic, Indo-Malayan, and Indo-Bhutan biogeographical realms, is a UNESCO World Heritage Site, a tiger reserve, an elephant reserv biosphere reserve, and a biodiversity hotspot. Most of the previous studies in MNP predominantly focused on aspects such as species diversity (Borah et al. 2012) and abundance (Borah et al. 2013; Borah et al. 2 Goswami & Ganesh 2014) and sympatric associations among large and medium-sized carnivores (Lahkar et al. 2021). Some exceptions, such as the research by Bhatt et al. (2021), have explored temporal interactions and l cycle influence on the entire terrestrial mammalian community. However, a critical gap exists, as no prior studies have investigated co-occurrence patterns and habitat use dynamics, specifically within the carnivore assemblage. The s region was ideal for examining such aspects, as it holds diverse carnivore guilds (17 species) with body sizes varying from large (>50 kg, three species), medium (>10-50 kg, two species), and small (< 10 kg, 12 species) (Bhatt et al. 2022).
Numerous studies have elucidated distinct spatial and habitat utilisation patterns among carni species (Grassman et al. 2005; Davis et al. 2011; Gompper et al. 2016; Chutipong et al. 2017; Chatterjee et al. 2020; Vernes et al. 2022). In this study, we employed camera traps to explore co-occurrence patterns (positive, negative, and random) within carnivore communities, encompassing large, medium, and small species, in the tropical forest of MNP, situated in northeast India, a vital abode in South Asia. Additionally, we examined the influence of various factors, such as habitat characteristics, anthropogenic variables, food abundance (bottom-up effects), and predation risk (top-down effects), on the habitat usage of carnivores. Our specific objectives were threefold: (i) to assess the composition of the carnivore community, (ii) to analyse co-occurrence patterns among carnivore species, and (iii) to identify the key factors driving habitat selection for each carnivore species. The information derived from this study cotributes to a deeper understanding of carnivore dynamics within tropical forest systems and holds significant potential in developing more effective management and conservation measures.
The primary aim of this study was to investigate if there was any spatial avoidance among sympatric carnivores captured at the same camera sites and if the detection of subdominant species was related to the detection of dominant species. Accordingly, we hypothesised random or negative co-occurrence between sympatric carnivores. Specifically, we predicted mesopredators, such as dholes and clouded leopards, would be less likely detected at camera sites where larger predators like tigers and leopards had been recorded. Moreover, we anticipated small carnivores would show reduced detection frequencies in areas occupied by mesopredators.
Six hypotheses were tested for carnivore habitat use: (1) As habitat degradation, agricultural expansion, overhunting, and poaching threaten Asian carnivores (Ashrafzadeh et al. 2020; Marneweck et al. 2021), we hypothesised a negative association between carnivore occurrence and anthropogenic variables (human settlements and roads). (2) As Human-induced large-scale fragmentation negatively affects the canopy cover and vegetation, which is a proxy for forage biomass and the distribution of large carnivores, and small forest patches provide refuge for small carnivores (Pettorelli 2013; Mallegowda et al. 2015), we hypothesised a positive relationship between carnivore species and forest cover (NDVI and canopy cover). We also anticipated the impact of forest cover on carnivore species would vary based on their specific ecological requirements. (3) As the elevation of the MNP, i.e., 40-200 m asl, is not a proximal factor but a distal factor that likely influences carnivore habitat use, we hypothesised that elevation would not affect carnivore species occurrence. (4) As resources (prey availability) are crucial determinants of carnivore habitat use (Chutipong et al. 2017), we hypothesised a positive relationship between the occurrence of large and medium-sized carnivores and prey availability. (5) As interspecific killing risks or predation risk effects (dominant predators) influence carnivore habitat use (Chutipong et al. 2017), we hypothesised a negative association of carnivore species with predators dominant to them. (6) As the risk effect from mesopredators potentially weakens in the presence of apex predators (Steinmetz et al. 2013), we hypothesised a positive relationship between small and large carnivores.
Through testing these hypotheses, the study aimed to shed light on the spatial dynamics and potential interactions between different carnivore species and their habitat use in the context of habitat variables, anthropogenic pressures, and ecological factors (such as prey availability and predation risk). These findings have valuable implications for the conservation and management of carnivores in the study area and beyond, as they provide insights into the critical threats these species face and offer guidance for developing effective conservation measures. Furthermore, these insights deepen our understanding of the co-existence of carnivores.
Material and Methods
Study area:
We conducted the study in an area of 500 km² within MNP (26°80’-26°50’ N, 90°45’-91°15’ E), a UNESCO World Heritage Site, in the state of Assam, India. MNP lies on the borders of the Indo-Gangetic, Indo-Malayan, and Indo-Bhutanbiogeographical realms on a gentle alluvial slope in the foothills of the Himalayas, where wooded hills give way to grasslands and tropical forests. The climate exhibits four seasons: (1) Pre-Monsoon: March-May, (2) Monsoon: June-September,(3) Retreating Monsoon: October-November and (4) Winter: December-February. The mean temperature ranges from 5-37 °C. The elevation ranges from 40-200 m asl (Bhattacharjya et al. 2015). The Park is home to a variety of important mammal species, including the tiger Panthera tigris, pygmy hog Porcula salvania, hispid hare Caprolagus hispidus, and Asiatic elephant Elephas maximus (Wikramanayake et al., 2002). Also, it supports22 of India’s most threatened mammal species, as listed in Schedule-I of the Wildlife (Protection) Act of India (DebRoy, 1991). Together with the Royal Manas National Park in Bhutan, the Park forms one of the largest areas forconservation significance in South Asia, representing the full range of habitats from the subtropical plains to the alpine zone (Wang, 2001). The habitat in the area was mainly intact and had only been mildly impacted by some cattlegrazing and the sustainable collection of firewood, timber, and non-wood forest products by local people at the edges of the National Park.
Field sampling design:
We used camera traps to photograph carnivores and prey species within strictly forested habitats of MNP. We selected camera trap locations based on accessibility, terrain features, animal trails, and nallahs (seasonal drainages) with carnivore
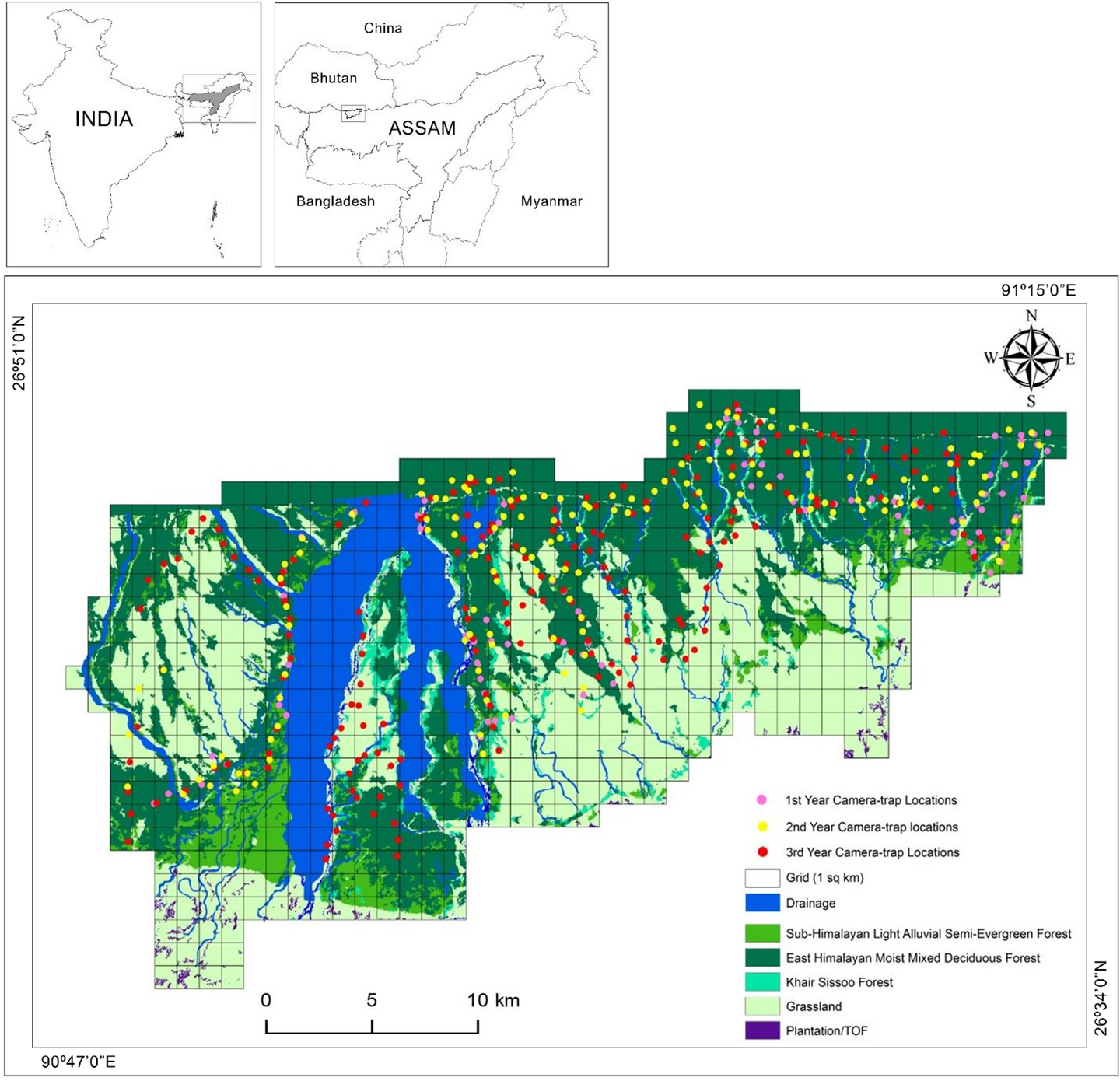
Figure 1: The study area (Manas National Park) map shows camera trap locations (n=473), drainage, and forest cover. Camera traps were deployed in a grid-based approach (grid size: 1 km²) during three sampling periods: April – June 2017 (n=101), December 2017–May 2018 (n=152), and November 2018 – May 2019 (n=220).
signs (Marinho et al., 2018). At each location, we set up a single Cuddeback X-Change™ colour model (Cuddeback, Green Bay, WI, USA) with motion sensors and a time lag of 1s between animal detections. We fastened the camerasto trees approximately 30-45 cm above the ground for 24 days on average (Range: 16-45). We utilised 473 camera trap locations in a grid-based approach with a grid size of 1 km² during three sampling periods: April 2017-June 2017 (n=101),December 2017-May 2018 (n=152), and November 2018-May 2019 (n=220) (Figure 1). We chose these months explicitly for greater accessibility and minimal variance in weather conditions. To optimise capture probability, we maintained amean distance of 672 meters (SE ± 15.73) between neighbouring camera traps. Previous studies have shown that the small Indian mongoose, the region’s smallest carnivore, has a home range of 0.450 km² (Berentsen et al., 2020).Therefore, we believe this inter-trap distance adequately covered the home ranges of all carnivores in our study area. The average temperature and humidity during the entire sampling period were 24⁰C (SE ± 0.29) and 79.4% (SE ± 0.54), respectively (Time and Date 2021).
Species richness, composition, and relative abundance index:
We estimated species richness as the total number of species detected during the study period. We obtained species-specific body masses from the literature and calculated them as the average male and female adult body masses. We classified mammals into three body-size classes: large (>50 kg), medium (>10-50 kg), and small ( < 10 kg) (Prater & Barruel, 1971; Menon, 2003; Wilson & Mittermeier, 2011; Menon, 2014). We calculated the relative abundance index (RAI) as the total number of independent photographs for each species divided by the total trap nights and multiplied the result by 100 (Carbone et al. 2001). We considered animal detections independent if the time between consecutive photographs of the same species was > 30 min (O’Brien et al., 2003). We conducted the analysis using Windows based MS Office Excel.
Carnivore co-occurrence and interactions:
We used a statistical co-occurrence test to evaluate the spatial interactions among carnivores. We created capture histories using daily capture events to determine the presence or absence of targeted species at each camera station. Using these capture histories, we investigated spatial interactions (positive, negative or random) between species pairs using probabilistic models (Veech, 2013; Griffith et al. 2016). We based these models on two probabilities (Veech, 2013). (1) Probability of lower co-occurrence frequency (P lt): This probability represents the likelihood that two species co-occur at a frequency lower than what was observed in the data. It indicates whether the observed co-occurrence between the species is less common than expected by chance. (2) Probability of higher co-occurrence frequency (P gt): This probability represents the likelihood that two species co-occur at a frequency higher than what was observed in the data. It indicates whether the observed co-occurrence between the species is more common than expected by chance. P lt < 0.05 and P gt < 0.05 indicate negative co-occurrence (competitive interaction) and positive co-occurrence (no antagonism) between the considered species pairs, respectively. Conversely, when P lt > 0.05 and P gt > 0.05, co-occurrence is considered genuinely random, implying an independent distribution of the species (Veech, 2013; Griffith et al., 2016).
We also determined the standardised effect sizes (range −1 to 1) as the difference between the observed and expected co-occurrence frequencies divided by the number of sampling points (Veech 2013). Additionally, we compared the relative abundances of carnivores to investigate spatial overlap (O’Brien et al. 2003). We considered each camera-trap site as spatially independent and used the relative abundance for each camera site to calculate the spatial overlap index (range 0–1; 0=no overlap; 1=complete overlap of the niche category by the species).
Carnivore habitat use
Habitat and anthropogenic variables:
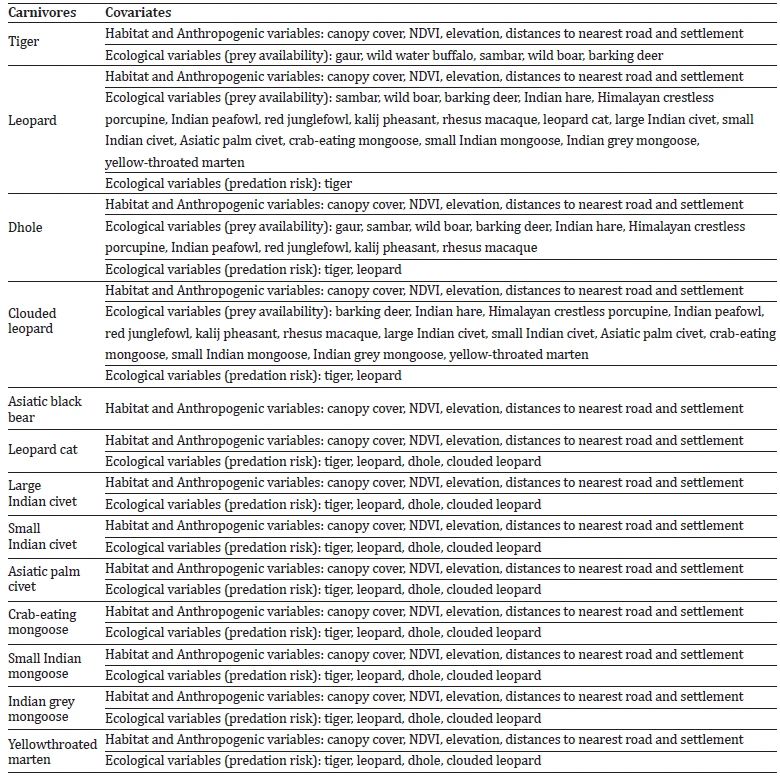
We focused on three habitats and two anthropogenic variables for carnivore habitat use: canopy cover, elevation, normalised difference vegetation index (NDVI), distance to the nearest settlement, and distance to the nearest road. We quantified tree canopy cover (30m resolution) using global tree cover data (Hansen et al., 2013). We determined the elevation at each camera trap site using a digital elevation model based on NASA’s ASTER Global Emissivity Datasets (USGS Earth Explorer 2021), with a spatial resolution of 30m. For NDVI, we downloaded Landsat images (30m resolution) from the Landsat 8 Operational Land Imager (OLI) sensor in bands 4 and 5. We then computed the NDVI as (𝑏𝑎𝑛𝑑 5 − 𝑏𝑎𝑛𝑑 4) / (𝑏𝑎𝑛𝑑 5 + 𝑏𝑎𝑛𝑑 4) (USGS Earth Explorer 2021). We quantified distances to the nearest settlement and road for each camera trap site using Euclidean distance and performed these calculations using ArcMap v. 10.2 (Esri, 2011).
Ecological variables:
We comprehensively reviewed the literature on carnivore diets to identify potential prey species in the study area (Supplementary Table S1). Our camera traps detected these or related species of this size range that are likely to be prey. We associated the presence of carnivore species with the relative abundance of prey species of a certain size class (i.e., large: >50 kg, medium: >10-50 kg, small: 1-10 kg) and based on carnivore diet literature (Supplementary Table S1). However, we excluded prey species weighing < 1 kg and arboreal mammals from the analysis. We decided to exclude these species because camera trapping studies indicated their low detection probability, which would not accurately represent their actual abundance (Tobler et al. 2008). Consequently, we could not quantify the prey species available for small carnivores. Furthermore, to investigate predation risk effects, we focused on understanding how dominant predators influence the habitat use of mesopredators and how both dominant and medium-sized predators affect the habitat use of lesser carnivores.
Data analysis:
We investigated the habitat use of carnivores by creating generalised linear models (GLMs) (Chatterjee et al., 2020; Chaudhary et al., 2020), using camera trap station-specific carnivore presence or absence (1,0) data, along with continuous covariates (habitat, anthropogenic, and ecological) (Table 1). To model carnivores’ habitat use, we standardised all continuous covariates using a z-transformation and reduced multicollinearity by removing highly correlated predictor variables (Spearman rank correlation |r 2|>0.7) (Nath et al., 2019). We fitted GLMs with a binomial link function and selected the model with the lowest Akaike information criterion (AIC) value and the highest Akaike model weights as the best-supported model (Anderson 2008). We predicted the potential habitat of carnivores using the coefficients from the best-fit model with the lowest AIC (Hijmans et al., 2013).
We performed all statistical analyses in R software v. 3.1.2 using packages MuMIn (Barton, 2016), co-occur (Griffith et al. , 2016), spaa (Zhang et al., 2013), and raster (Hijmans et al., 2013).
Results
Species richness, composition, and relative abundance index (RAI):
We recorded 37 species (17 carnivores and 20 non-carnivores), with 24,840 independent records over the entire sampling period of 11,388 trap nights (Supplementary Table S2). Independent records (n) and RAI for the photo-captured species varied, ranging from clouded leopard (n=21, RAI=0.18) to tiger (n=466, RAI=4.09) for large-medium carnivores, from Chinese pangolin (n=2, RAI=0.02) to small Indian civet (n=402, RAI=3.53) for small carnivores, from chital (n=1, RAI=0.01) and Himalayan goral (n=1, RAI=0.01) to Asiatic
Table 1: Selection of covariates (habitat, anthropogenic, and ecological) employed in generalised linear modelling of carnivores in Manas National Park.
elephant (n=6386, RAI=56.08) for large-medium herbivores, and from Asiatic brush-tailed porcupine (n=3, RAI=0.03) to red junglefowl (n=1162, RAI=10.20) for small herbivores. We excluded species with less than ten detections and habitat specialist species from the analysis.
Carnivore co-occurrence and interactions:
We analysed 78 species pairs of 13 carnivores. We excluded ten pairs (12.82% of the 78 combinations) from the analysis because the expected co-occurrence for these pairs was less than one. Three species pairs in the large-medium carnivore guild (tiger–leopard, tiger–dhole, leopard–dhole), eleven in the small carnivore guild (leopard cat–large Indian civet, leopard cat–small Indian civet, leopard cat–Asiatic palm civet, leopard cat–crab-eating mongoose, large Indian civet–small Indian civet, large Indian civet–crab-eating mongoose, small Indian civet–Asiatic palm civet, small Indian civet–small Indian mongoose, small Indian civet–crabeating mongoose, small Indian civet–yellow-throated marten, Asiatic palm civet–crab-eating mongoose), and five in the inter-guild (tiger–leopard cat, tiger–large Indian civet, leopard–leopard cat, leopard–large Indian civet, clouded leopard–leopard cat) showed positive co-occurrences (Figure 2). The Pianka’s index values between these pairs were low, with the highest value reported between Asiatic palm civet and crab-eating mongoose, which was 0.392 (0.239-0.535) (Table S3). We did not observe clear co-occurrence patterns for the remaining 59 species pairs, indicating the random distribution of these species pairs across the study area.
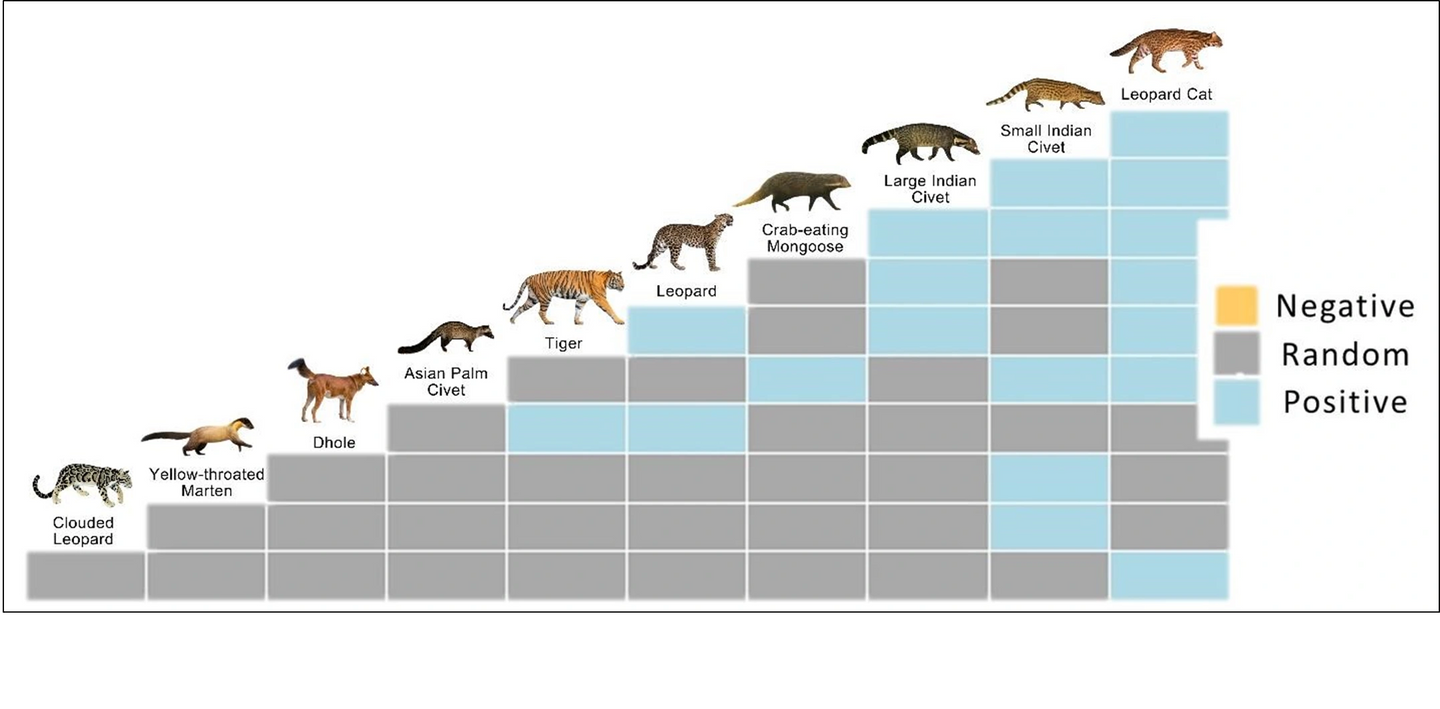 Figure 2: Heat map shows the positive, random, and negative species associations determined by the probabilistic co-occurrence model for carnivores in Manas National Park. Species are positioned to indicate the columns and rows that represent their pairwise relationships with other species. 78 pairs were analysed, of which 19 were positive (no antagonism), while 59 showed random (independent) distribution.
Figure 2: Heat map shows the positive, random, and negative species associations determined by the probabilistic co-occurrence model for carnivores in Manas National Park. Species are positioned to indicate the columns and rows that represent their pairwise relationships with other species. 78 pairs were analysed, of which 19 were positive (no antagonism), while 59 showed random (independent) distribution.
Carnivore habitat use
Large-medium carnivores:
The analysis of habitat use patterns through GLMs revealed distinct habitat preferences among carnivore species (Table 2, Supplementary Table S4). While examining predictor variables, we found no significant relationships; therefore, we retained all variables for further analysis. For tiger, habitat use intensity showed positive associations with distance to roads, the presence of gaur, wild water buffalo, sambar, and wild boar, and a negative association with elevation. In the case of leopard, predictor variables included tiger, wild boar, red junglefowl, and leopard cat. Dhole habitat utilisation showed positive associations with tiger and rhesus macaque and a negative association with kalij pheasant. Clouded leopard showed higher habitat use in areas with more prevalent canopy cover, NDVI, red junglefowl, and kalij pheasant. Asiatic black bear habitat use exhibited a positive relationship with elevation and a negative association with distances to roads and settlements.
Three models exhibited the best performance for tiger and leopard, while two for dhole, clouded leopard, and Asiatic black bear with ΔAIC < 2 (Table 3, Supplementary Table S5).
Small carnivores:
Canopy cover and NDVI influenced the habitat utilisation of leopard cat, Asiatic palm civet, and yellow-throated marten (Table 2, Supplementary Table S6). The habitat use of large Indian civet and Asiatic palm civet showed positive associations with elevation. Leopard cat, large Indian civet, Asiatic palm civet, and yellow-throated marten exhibited a preference for habitats situated farther away from human settlements. Crab-eating mongoose habitat showed a positive relationship with the proximity of human settlements.
The analysis produced nine best-performing models for yellow-throated marten, five for Asiatic palm civet, four for each large Indian civet and small Indian civet, three for leopard cat, two for Indian grey mongoose, and one for each crab-eating mongoose and small Indian mongoose, all with ΔAIC < 2 (Table 3, Supplementary Table S7).
We then generated spatial prediction maps for large, medium, and small carnivores using the best-fit model with the lowest AIC and highest weights (Figures 3, 4).
Discussion
We conducted a comprehensive camera trap survey in MNP to document the community of small, medium, and large mammalian carnivores and understand their habitat associations. Our fine-scale modelling approach incorporated various variables, including habitat, anthropogenic influence, and ecological factors, revealing species-specific environmental responses. These findings reaffirmed our knowedge about carnivores and provided valuable insights into their habitat preferences. By analysing co-occurrence patterns and environmental drivers, this research offers crucial information for carnivore conservation and management, contributing significantly to our understanding of their ecology and behaviour in MNP.
As hypothesised, negative or random co-occurrences were expected between intra- and inter-guild carnivores. However, a few pairs (n=19) revealed positive co-occurrences, while most pairs (n=59) showed random cooccurrences or an independent distribution (Rota et al., 2016), partially supporting the spatial avoidance hypothesis. This suggests that species with positive co-occurrence must be differentiated along other niche dimensions, such as time and diet, to facilitate their co-existence. For instance, Bhatt et al., (2021) found evidence that these carnivore species avoided each other by displaying low temporal overlap or having distinct activity peaks. Additionally, variations in their diet patterns (e.g., carnivore, omnivore, and insectivore) further contribute to species co-occurrence by enabling niche partitioning on a microhabitat scale. However, further research is required to identify the specific variables governing their co-occurrence and potential interactions at this fine-scale level.
Table 2: Species-specific significant beta coefficients with standard errors, Z-scores, and P-values of habitat, anthropogenic, and ecological variables influencing habitat use of (a) large-medium carnivores and (b) small carnivores in Manas National Park. Beta coefficients represent the strength and direction of the relationship between variables and carnivore habitat use. Standard errors provide an estimate of the variability in the beta coefficients. Z-scores indicate the number of standard deviations a beta coefficient is from the mean. P-values determine the statistical significance of the beta coefficients, with lower values indicating stronger evidence against the null hypothesis. Significance levels: ** p < 0.05, * p < 0.10 (where ** denotes high significance and * denotes moderate significance).
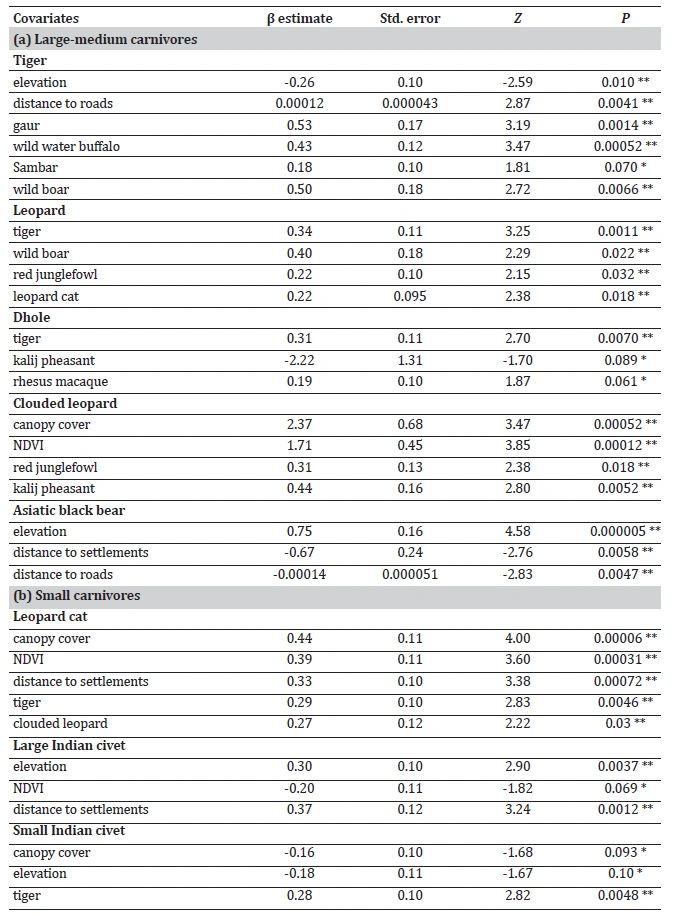
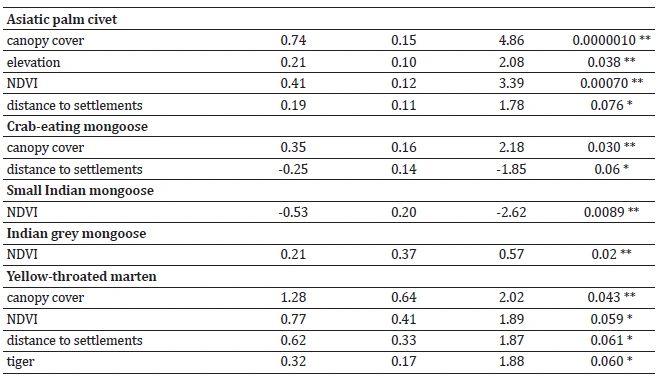
Table 3: Best-fit generalised linear models for assessment of habitat use of (a) large-medium carnivores and (b) small carnivores in Manas National Park. The models were selected based on their goodness-of-fit statistics, with ΔAIC < 2 indicating strong support for model fit
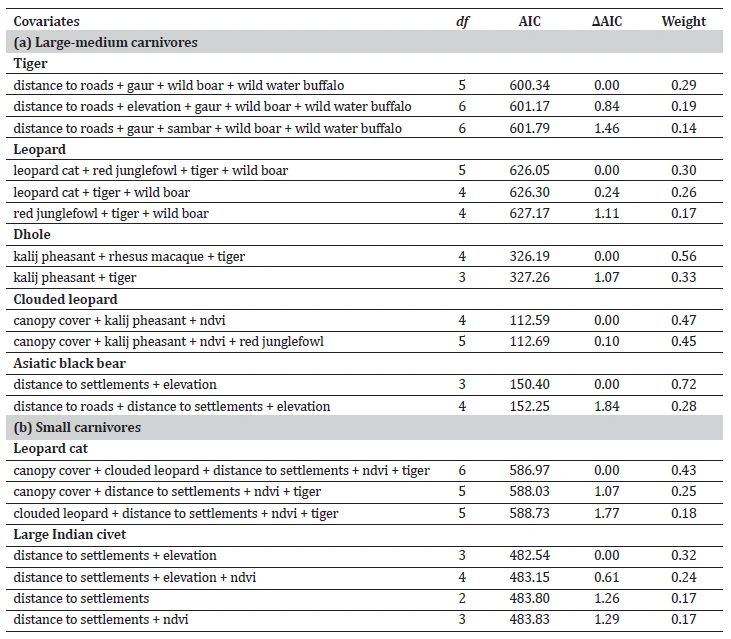
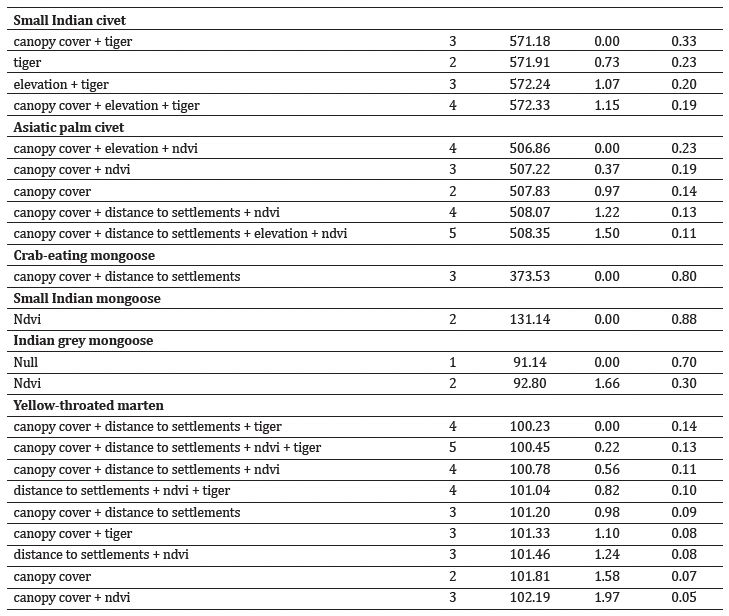
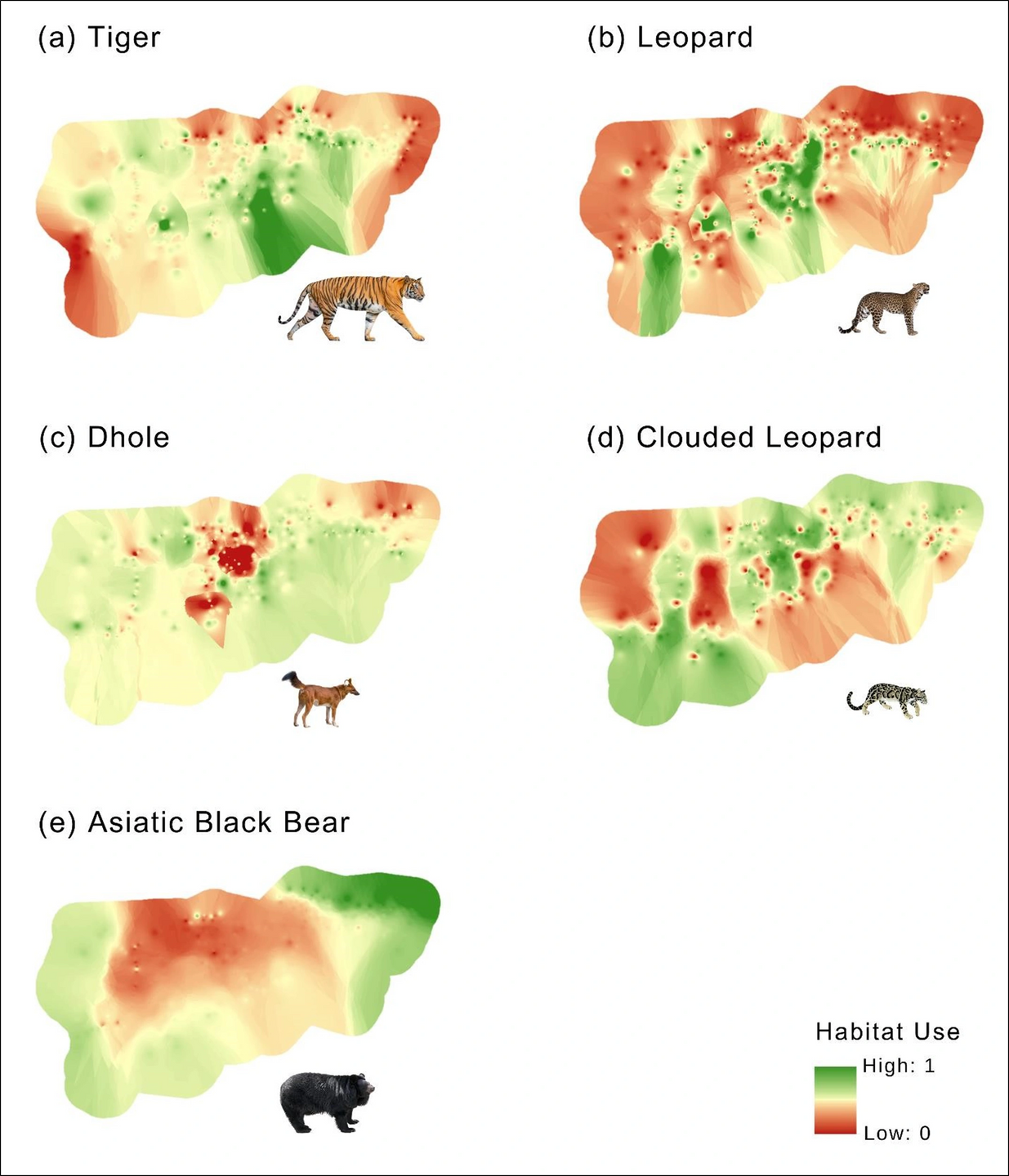
Figure 3: Spatial prediction for potential habitat use of large-medium carnivores using best-fit model with lowest AIC and highest model weight in Manas National Park. (a) Tiger: distance to road + gaur + wild water buffalo + wild boar, (b) Leopard: tiger + wild boar + red junglefowl + leopard cat, (c) Dhole: tiger + rhesus macaque + kalij pheasant, (d) Clouded leopard: canopy cover + NDVI + kalij pheasant, and (e) Asiatic black bear: elevation + distance to settlements.
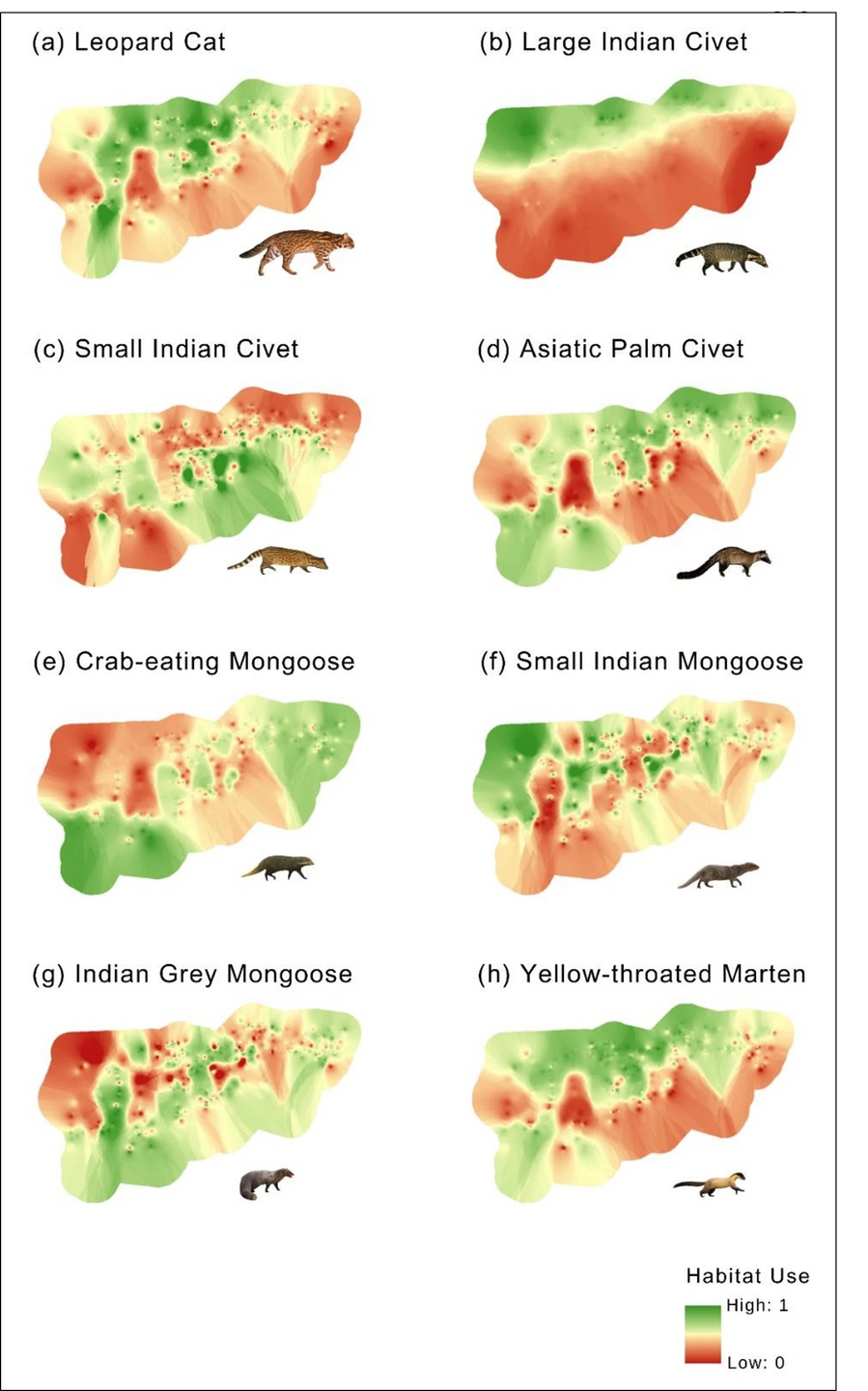
Figure 4: Spatial prediction for potential habitat use of small carnivores using best fit model with lowest AIC and highest model weight in Manas National Park. (a) Leopard cat: canopy cover + NDVI + distance to settlements + clouded leopard + tiger, (b) Large Indian civet: elevation + distance to settlements, (c) small Indian civet: canopy cover + tiger, (d) Asiatic palm civet: canopy cover + NDVI + elevation, (e) Crab-eating mongoose: canopy cover + distance to settlements, (f) Small Indian mongoose: NDVI, (g) Indian grey mongoose: null, (h) Yellow-throated marten: canopy cover + distance to settlements + tiger.
Tiger, known for its adaptability across diverse landscapes (Goodrich et al., 2022), displayed no significant correlation with habitat variables except for elevation. Given the limited elevation range within MNP, elevation is unlikely to influence tiger habitat use significantly. Notably, the study revealed a negative impact of forest roads on tiger (Linkie et al., 2006). These roads, frequented by tourists, locals, and patrolling staff, pose challenges to tiger conservation efforts, necessitating careful management and monitoring to mitigate their adverse effects. In terms of prey preferences, tigers have a strong affinity for specific species, including wild boar, deer species, wild buffalo, gaur, elephant, rhino, primates, and porcupines (Supplementary Table S1). Intriguingly, the availability of these preferred prey species, such as gaur, wild water buffalo, sambar, and wild boar, appeared to influence tiger habitat usage. This highlights the role of prey availability in shaping the tiger’s habitat selection within the park.
Leopard, renowned for its adaptability to various landscapes (Stein et al., 2020), did not exhibit significant correlations with habitat variables, affirming its ability to thrive across diverse environments. Its diet mainly comprises of medium-sized prey (10-50 kg body mass) but also includes a diverse range of species, such as primates, rodents, lagomorphs, pangolins, small carnivores, gallinaceous birds, insects, reptiles, and birds (Table S1). The study also revealed that medium and small prey species influenced leopard habitat usage. In contrast to the expectations, a positive relationship was observed between leopard (subdominant predator) and tiger (dominant predator), suggesting a unique co-existence strategy. This association might be attributed to overlapping prey availability, habitat suitability, or the arboreal behaviours of leopard. The findings indicate the pivotal role of prey availability rather than predation risk in shaping leopard’s habitat preferences.
Asiatic black bear inhabits various forested habitats, ranging from near-sea level to 4,300 m in elevation (Sathyakumar et al., 2011; Garshelis & Steinmetz, 2020). Despite the narrow elevation range within the MNP, a positive relationship with elevation was observed (Sathyakumar et al., 2013). One reason could be that black bear may prefer higher elevations for denning to avoid potential predators. Another possible reason for its preference for higher elevations could be the earlier appearance of herb and grass sprouts in those areas than at lower elevations. Moreover, we found a positive association between Asiatic black bear and human settlements and roads. This could be attributed to bear’s learned tolerance for humans (human habituation) and its association of humans with readily available high-quality food (food conditioning) (Elfström et al., 2014).
Dholes, with a preference for various habitats and elevations (Kamler et al., 2015), displayed no significant relationship with habitat variables. Dhole diet encompasses a wide range of prey species, including small rodents, hares, and gaur (Selvan et al., 2013a, 2013b; Hayward et al., 2014). Their hunting success is influenced by various factors such as pack size, prey species, and habitat characteristics (Acharya, 2007). In tropical semi-evergreen forests, dholes persist in smaller packs and primarily consume small-to-medium-sized prey as they offer energetic advantages (Kawanishi & Sunquist, 2008). The findings of this study supported this pattern and suggested that the occurrence of dhole is influenced by small prey species, specifically rhesus macaque, highlighting the importance of conserving small sized species for the long-term viability of dhole. The negative relationship between dhole habitat use and pheasant could be attributed to habitat requirements and ecological preference differences. Pheasants are ground-dwelling birds that prefer dense undergrowth and shrubby areas (Miller, 2010), whereas dhole is a cursorial predator that requires more open spaces for hunting (Johnsingh, 1992). The contrasting habitat preferences of pheasants and dhole could have resulted in a negative association between them.
Clouded leopard has shown positive associations with NDVI and canopy cover (Austin et al., 2010; Brodie & Giordano, 2012; Penjor et al., 2018), and our findings align with these findings. While previous research suggested a preference for high elevations (Ngoprasert et al., 2012; Haidir et al., 2013; Mohamad et al., 2015), we found that the elevation of MNP is a distal factor that likely influences its habitat use. Grassman et al. (2005) speculated, based on their radio telemetry data, that clouded leopard hunts smaller prey (≤2.5 kg) and assumed it would move less if it killed larger prey. The study supported this assumption, suggesting that prey species of < 10 kg body mass, specifically galliformes (kalij pheasant, 1.1 kg; red junglefowl, 1.25 kg; Indian peafowl, 5.0 kg), had a significant influence on clouded leopard occurrence. However, a study in 13 protected areas in Thailand showed that clouded leopards is associated with wild boar and southern red muntjac (Ngoprasert et al., 2012); it did not test for the effect of smaller potential prey species.
Our findings suggest that conserving small-sized species such as primates, porcupines, hares, and galliformes is critical to increasing the viability of mesopredators (dhole and clouded leopard). In addition, high tiger and leopard densities in MNP (Borah et al., 2014; Supplementary Table S8) increase competition for larger prey and could further increase the relative importance of smaller prey for both medium-sized carnivores. However, dietary studies are required to examine whether these species form a part of their diet. Our prediction that medium-sized predators avoid larger apex predators behaviourally is based on the notion that the local distribution of a top predator is determined by resource availability, but the distribution of a mesopredator is influenced primarily by predation risk (Thompson & Gese, 2007). Although we did not detect a substantial influence of large predator on the habitat use of clouded leopard, our findings found a positive relationship between dhole and tiger. The positive association could be attributed to factors such as overlapping prey availability or habitat suitability, which may facilitate co-existence or even cooperative behaviour between dhole and tiger. Moreover, the findings suggest that dhole and clouded leopard choose habitats based on high prey abundance.
Leopard cat, typically considered ground-dwelling species, has been observed resting high in the canopy (Rabinowitz, 1990). Our research showing a positive association between leopard cats and canopy cover and NDVI aligns with this observation. Affinity for higher canopy and denser vegetation indicates the preference for areas providing suitable resting and hunting grounds. Large Indian civet displays remarkable habitat adaptability, occupying a wide range of environments, from primary forests to secondary degraded forests, scrublands, and plantations (Azlan, 2003; Jennings and Veron, 2011; Choudhury, 2013; Chutipong et al., 2014). This diverse habitat use explains their antagonistic relationship with ndvi, potentially resulting in lower occurrences in areas with dense vegetation. Small Indian civets, conversely, favours low elevations and open habitats (Jennings & Veron, 2011), avoiding areas with dense vegetation or higher elevations, reflecting their adaptability to varying habitat conditions. Asiatic palm civet inhabits a diverse range of habitats, spanning evergreen and deciduous forests, seasonal peat swamp forests, mangroves, plantations, villages, urban areas, and elevations up to 2500 m (Duckworth et al., 2016). However, our study revealed a positive association between Asiatic palm civet site preferences and the presence of primary forests. This preference may be attributed to a rich and diverse food supply, enhanced shelter and security, and suitable conditions for breeding and reproduction.
Crab-eating mongoose, Indian grey mongoose, and Yellow-throated marten preferred habitats characterised by dense vegetation. Such vegetation provides these small carnivores suitable shelter, cover, and protection from predators. Dense vegetation also facilitates a favourable environment for hunting and foraging, as it harbours various prey species and provides ample hiding spots for these ambush predators.
Small felids, civets, and martens are hunted for their meat, providing a vital protein source for numerous local tribal communities in tropical regions (Corbet & Hill, 1992). The study also revealed a negative relationship between humans and leopard cats, large Indian civets, Asiatic palm civets, and yellow-throated martens, indicating potential threats to these species from human activities. On the other hand, mongooses have a commensal relationship with humans, as they benefit from scavenging on carrion and human waste near human habitations (Choudhury et al., 2011). These findings align with the expected pattern, mainly for crab-eating mongoose.
The findings of this study demonstrated a positive association between specific small carnivores, such as leopard cat, small Indian civet, and yellow-throated marten, and the presence of large carnivores, particularly tiger. Large carnivores play a crucial role as apex predators, effectively controlling meso predators, which benefits smaller carnivores. This top-down regulation within the carnivore community helps maintain a balanced ecosystem by reducing predation and competition risks for smaller species, thereby promoting their thriving. Consequently, conservation efforts aimed at protecting and conserving large carnivores will have positive implications for smaller ones and contribute to maintaining the overall ecological balance of the ecosystem.
Regarding the interpretation of P-values, we acknowledge the need to consider the broader context and effect sizes rather than relying solely on arbitrary thresholds (Wasserstein et al., 2019). Although the P-values for a few variables indicate a lack of statistical significance according to traditional criteria, we emphasise that these marginal results offer valuable insights and warrant further investigation or consideration in future studies. The selection of the best fit model for prediction purposes allowed us to gain valuable insights into the factors influencing carnivore habitat use and provided a spatial representation of their potential habitat within MNP. This spatial mapping offers a valuable tool for conservation planning and management efforts, as it highlights the areas where carnivores are likely to thrive and where conservation actions can be targeted to ensure the long-term survival of this species.
The diet of the Asiatic black bear and small carnivores encompasses a diverse range of prey items, including fruits (cinnamon, pea, etc.), succulent vegetation (shoots, forbs, and leaves), ants, bees, a variety of tree and shrub-borne fruits, hard mast (nuts), rodents, birds, murids, fishes, carrion, reptiles, amphibians, insects, arthropods, and crabs (Supplementary Table S1). However, we could not analyse these prey species as camera trapping rates would not reflect their abundance due to low detection probability (Ancrenaz et al., 2012). Therefore, habitat use for the prey availability of these carnivorous species cannot be estimated.
Conservation, both political lobbying and on-the-ground management, can only be effective with full knowledge of which species are present in an area and their distributions and habitat use. Manas National Park was affected by unrest that occurred in the region from the mid-1980s until peace was restored in 2003 (Goswami & Ganesh, 2014), leaving it vulnerable to logging, hunting and poaching of fauna and resulting in habitat degradation and significant loss of wildlife (George, 1994; Sarma et al., 2008). Nevertheless, our study provides evidence that most carnivore species in Manas use habitats rich in primary forests, abundant prey and far from human disturbances. The observed relationships between carnivore species and habitat characteristics are likely shaped by a complex interplay of factors, including food availability, shelter, competition avoidance, and species-specific behaviour and adaptability. Understanding these relationships is crucial for monitoring conservation success, planning conservation intervention plans, and implementing effective conservation strategies to ensure the long-term survival of carnivores and their coexistence in their habitats.
Conservation efforts in Manas National Park should prioritise protecting and conserving primary forest habitats, which are crucial refuges for many carnivore species due to their rich biodiversity and ample prey availability. Measures to mitigate human disturbances, such as managing forest roads and minimising human-wildlife conflicts, are vital for reducing habitat fragmentation, which can adversely affect carnivore populations. Moreover, promoting co-existence between humans and carnivores through community-based conservation initiatives and sustainable land-use practices is essential. Finally, it is critical to maintain healthy prey populations within the Park, as they form the foundation of the carnivore food chain and play a vital role in sustaining carnivore populations. Together, these conservation recommendations can contribute to the long-term conservation success of carnivores in Manas National Park.
TO DOWNLOAD SUPPLEMENTARY MATERIAL CLICK HERE
Acknowledgement
We thank the Director and Dean of the Wildlife Institute of India for their guidance and support. We are grateful to (Lt) Doyil Vengayil, Syed Asrafuzzaman, and Harish Kumar, Department of Science and Technology, Government of India, for financial assistance to the project on clouded leopards under grant no. EMR/2015/000085 of 01-04-2016. We thank the Forest Department, Government of Assam, and Bodoland Territorial Council (BTC) for permitting us to conduct the survey. We are grateful to Mr Hiranya Kumar Sarma (IFS, former Field Director, MTR), Mr Amal Chandra Sarma (IFS, former Field Director, MTR), Dr Vaibhav Chandra Mathur (IFS, Field Director, MTR), and Mr Abbas Dewan (ACF, MTR) for facilitating to work in MNP. We thank the range officers of the MNP, Babul Brahma, Kunjan Basumatary, Kameshwar Boro, (Lt) Pranab Das and the frontline staff for providing logistic support during the field surveys. We thank our field assistants and drivers (Paniram, Dipen, Dipul, Umesh, Bablu, Suraj, Bhadreshwar, and Kangkan). Thanks to Paniram, Tejas, Chiging, Meban, Saurav, Nikunj, and Krishna for assisting in the field. We appreciate our reviewers’ and editors’ valuable comments and suggestions for improving the manuscript.
CONFLICT OF INTEREST
The authors have no competing interests to declare that are relevant to the content of this article.
DATA AVAILABILITY
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
AUTHORS’ CONTRIBUTION
SL, UB conceptualised the study. UB collected data. UB, SL curated data. SL, UB developed methodology. UB, SL analysed data. UB wrote the original manuscript. SL, UB reviewed and edited the manuscript. SL procured funding, provided resources, and supervised the work.
Edited By
Ajith Kumar
National Centre for Biological Sciences
*CORRESPONDENCE
Salvador Lyngdoh
✉ salvador@wii.gov.in
CITATION
Bhatt, U., Lyngdoh, S. (2024). Carnivore Chronicles: Co-occurrence and habitat use in the tropical forest of Manas National Park, North-East India. Journal of Wildlife Science,1(1), 16-30. https://doi.org/10.63033/JWLS.GCTQ1198
FUNDING
The study was funded by Department of Science and Technology, Govt. of India under grant no. EMR/2015/000085 of 01-04-2016.
COPYRIGHT
© 2024 Bhatt, Lyngdoh. This is an open-access article, immediately and freely available to read, download, and share. The information contained in tarticle is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0![]()
![]() ), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepacademic practice. Copyright is retained by the author(s).
), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepacademic practice. Copyright is retained by the author(s).
PUBLISHED BY
Wildlife Institute of India, Dehradun, 248 001 INDIA
PUBLISHER'S NOTE
The Publisher, Journal of Wildlife Science or Editors cannot be held responsible for any errors or consequences arising from the use of the informatcontained in this article. All claims expressed in this article are solely those of the author(s) and do not necessarily represent those of their affiliated organisations or those of the publisher, the editors and the reviewers. Any prodthat may be evaluated or used in this article or claim made by its manufacturer is not guaranteed or endorsed by the publisher.
Acharya, B. B. (2007). The Ecology of the Dhole or Asiatic Wild Dog (Cuon alpinus) in Pench Tiger Reserve, Madhya Pradesh. PhD dissertation, Saurashtra University, Rajkot, India.
Ancrenaz, M., Hearn, A. J., Ross, J., Sollmann, R. & Wilting, A. (2012). Handbook for Wildlife Monitoring Using Camera-Traps. BBEC Publication, Sabah, Malaysia, 71.
Anderson, D. R. (2008). Model Based Inference in the Life Sciences. Springer, New York, 184. https://doi.org/10.1007/978-0-387-74075-1
Araujo, M. B. & Guisan, A. (2006). Five (or so) challenges for species distribution modelling. Journal of Biogeography, 33(10), 1677–1688. https://doi.org/10.1111/j.1365-2699.2006.01584.x
Ashrafzadeh, M. R., Khosravi, R., Adibi, M. A., Taktehrani, A., Wan, H. Y. & Cushman, S. A. (2020). A multi-scale, multi-species approach for assessing effectiveness of habitat and connectivity conservation for endangered felids. Biological Conservation, 245, 108523. https://doi.org/10.1016/j.biocon.2020.108523
Austin, S. C., Tewes, M. E., Grassman, Jr. L. I. & Silvy, N. J. (2010). Ecology and conservation of the leopard cat Prionailurus bengalensis and clouded leopard Neofelis nebulosa in Khao Yai National Park, Thailand. Acta Zool Sinica, 53, 1–14.
Azlan, J. (2003). The diversity and conservation of mustelids, viverrids, and herpestids in a disturbed forest in Peninsular Malaysia. Small Carnivore Conservation, 29, 8–9.
Barton, K. (2016). MuMIn: Multi-Model Inference. R package version, 1.47.1.
Berentsen, A. R., Rivera-Rodriguez, M. J., McClure, K. M., Torres-Toledo, F. B., Garcia-Cancel, J. G. & Gilbert A. T. (2020). Home range estimates for small Indian mongooses (Urva auropunctata) in southwestern Puerto Rico. Caribbean Journal of Science, 50(2), 225–235. https://doi.org/10.18475/cjos.v50i2.a4
Bhatt, U., Adhikari, B. S., Habib, B. & Lyngdoh, S. (2021). Temporal interactions and moon illumination effect on mammals in a tropical semievergreen forest of Manas National Park, Assam, India. Biotropica, 53(3), 831–845. https://doi.org/10.1111/btp.12917
Bhatt, U., Adhikari, B. S. & Lyngdoh, S. (2022). Monitoring diversity and abundance of mammals with camera-traps: a case study of Manas National Park, Assam, India. Check List, 18(5), 1023–1043. https://doi.org/10.15560/18.5.1023
Bhatt, U. & Lyngdoh, S. (2023a). Secrets of the clouded leopard: abundance, habitat use and carnivore coexistence in tropical forest of Manas National Park, Assam, India. Oryx, 57(6), 757–768. https://doi.org/10.1017/S0030605322001661
Bhatt, U. & Lyngdoh, S. (2023b). Do dholes segregate themselves from their sympatrids? Habitat use and carnivore co-existence in the tropical forest. Mammalian Biology, 103(6), 591–601. https://doi.org/10.1007/s42991-023-00378-z
Bhattacharjya, D. K., Kar, A., Sarma, H. & Patowary, K. N. (2015). Notes on herbal treatment practiced by the people of fringe villages of Manas National Park. India. Indian Journal of Traditional Knowledge, 1(1), 155–160.
Borah, J., Sharma, T., Das, D., Rabha, N., Kakati, N., Basumatary, A., Ahmed, M. F. & Vattakaven, J. (2014). Abundance and density estimates for common leopard Panthera pardus and clouded leopard Neofelis nebulosa in Manas National Park, Assam, India. Oryx, 48(1), 149–155. https://doi.org/10.1017/S0030605312000373
Borah, J., Sharma, T., Das N., Rabha, N., Kakati, N., Basumatri, A., Ahmed, F., Vattakaven, J., Bhobora, C. & Swargowari, A. (2012). Diversity of carnivores in Manas National Park - a World Heritage Site, Assam, India. Cat News, 56, 16–19.
Borah, J., Wangchuk, D., Swargowari, A., Wangchuk, T., Sharma, T., Das, D., Rabha, N., Basumatari A., Kakati N. et al. (2013). Tigers in the Transboundary Manas Conservation Complex: conservation implications across borders. Parks, 19, 51–62.
Brodie, J. & Giordano, A. J. (2012). Density of the Vulnerable Sunda clouded leopard Neofelis diardi in a protected area in Sabah, Malaysian Borneo. Oryx, 46(3), 427–430. https://doi.org/10.1017/S0030605312000087
Carbone, C., Christie, S., Conforti, K., Coulson, T., Franklin, N., Ginsberg, J. R., Griffiths, M., Holden, J., Kawanishi, K. et al. (2001) The use of photographic rates to estimate densities of tigers and other cryptic mammals. Animal Conservation, 4 (1), 75–79. https://doi.org/10.1017/S1367943001001081
Chatterjee, N., Nigam, P. & Habib, B. (2020). Population density and habitat use of two sympatric small cats in a central Indian reserve. Plos One, 15(6), e0233569. https://doi.org/10.1371/journal.pone.0233569
Chaudhary, R., Zehra, N., Musavi, A. & Khan, J. A. (2020). Evaluating the effect of ecological and anthropogenic variables on site use by sympatric large carnivores in Gir protected area, Gujarat, India. Wildlife Biology, 4, 1–7. https://doi.org/10.2981/wlb.00696
Choudhury, A. (2013). The mammals of North east India. Gibbon Books and the Rhino Foundation for Nature in NE India, Guwahati, Assam, India, 432.
Choudhury, A., Wozencraft, C., Muddapa, D., Yonzon, P., Jennings, A. & Geraldine, V. (2011). Herpestes edwardsii. In: IUCN 2012. IUCN Red List of Threatened Species, version 2012.2. www.iucnredlist.org. (Accessed on 03 November 2012).
Chutipong, W., Steinmetz, R., Savini, T. & Gale, G. A. (2017). Assessing resource and predator effects on habitat use of tropical small carnivores. Mammal Research, 62(1), 21–36. https://doi.org/10.1007/s13364-016-0283-z
Chutipong, W., Tantipisanuh, N., Ngoprasert, D., Lynam, A. J., Steinmetz, R., Jenks, K. E., Grassman, Jr. L. I, Tewes, M., Kitamura, S. & Baker, M. C. (2014). Current distribution and conservation status of small carnivores in Thailand: a baseline review. Small Carnivore Conservation, 51, 96–136.
Corbet, G. B. & Hill, J. E. (1992). The mammals of the Indo-Malayan region: a systematic review. Oxford University Press, Oxford, and Natural History Museum, London, 496.
Davis, M. L., Kelly, M. J. & Stauffer, D. F. (2011). Carnivore co-existence and habitat use in the Mountain Pine Ridge Forest Reserve, Belize. Animal Conservation, 14(1), 56–65. https://doi.org/10.1111/j.1469-1795.2010.00389.x
DebRoy, S. (1991). Manas: a monograph. Tiger paper (FAO), 18, 6–15.
Di Bitetti, M. S., De Angelo, C. D., Di Blanco, Y. E. & Paviolo, A. (2010). Niche partitioning and species co-existence in a Neotropical felid assemblage. Acta Oecologica, 36(4), 403– 412. https://doi.org/10.1016/j.actao.2010.04.001
Duckworth, J. W., Timmins, R. J., Choudhury, A., Chutipong, W., Willcox, D. H. A., Mudappa, D., Rahman H., Widmann, P., Wilting, A. & Xu, W. (2016). Paradoxurus hermaphroditus (Pallas, 1777). The IUCN Red List of Threatened Species 2016.
Elfström, M., Zedrosser, A., Støen, O. G. & Swenson, J. E. (2014). Ultimate and proximate mechanisms underlying the occurrence of bears close to human settlements: review and management implications. Mammal Review, 44(1), 5–18. https://doi.org/10.1111/j.1365-2907.2012.00223.x
ESRI (2011). ArcGIS Desktop: Release 10. Environmental System Research Institute, Inc., Redlands, California, USA.
Garshelis D. & Steinmetz R. (2020) Ursus thibetanus (amended version of 2016 assessment). The IUCN Red List of Threatened Species 2020: e.T22824A166528664.
George, S. J. (1994). The Bodo movement in Assam: Unrest to accord. Asian Survey, 34(10), 878–892. https://doi.org/10.2307/2644967
Glen, A. S. & Dickman, C. R. (2005). Complex interactions among mammalian carnivores in Australia, and their implications for wildlife management. Biological Reviews, 80(3), 387–401. https://doi.org/10.1017/S1464793105006718
Goldman, H. V. & Winther-Hansen, J. (2003). First photographs of the Zanzibar servaline Genet Genetta servalina archeri and other endemic subspecies on the island of Unguja, Tanzania. Small Carnivore Conservation, 29, 1–4.
Gompper, M. E., Lesmeister, D. B., Ray, J. C., Malcolm, J. R. & Kays, R. (2016). Differential Habitat Use or Intraguild Interactions: What Structures a Carnivore Community? Plos One, 11(1), e0146055. https://doi.org/10.1371/journal.pone.0146055
Goodrich, J., Wibisono, H., Miquelle, D., Lynam, A. J., Sanderson, E., Chapman, S., Gray, T. N. E., Chanchani, P. & Harihar, A. (2022). Panthera tigris. The IUCN Red List of Threatened Species, 2022, e.T15955A214862019.
Goswami R. & Ganesh, T. (2014). Carnivore and herbivore densities in the immediate aftermath of ethno-political conflict: the case of Manas National Park, India. Tropical Conservation Science, 7(3), 475–487. https://doi.org/10.1177/194008291400700308
Grassman, Jr. L. I, Tewes, M. E., Silvy, N. J. & Kreetiyutanont, K. (2005). Ecology of three sympatric felids in a mixed evergreen forest in north-central Thailand. Journal of Mammalogy, 86, 29–38. https://doi.org/10.1644/1545-1542(2005)086<0029:EOTSFI>2.0.CO;2
Griffith, D. M., Veech, J. A. & Marsh, C. J. (2016). Co-occur: Probabilistic species co-occurrence analysis in R. Journal of Statistical Software, 69(2), 1–17. https://doi.org/10.18637/jss.v069.c02
Griffiths, M. & Van Schaick, C. P. (1993). Camera-trapping: A new tool for the study of elusive rain forest animals. Tropical Biodiversity, 1, 131–135.
Haidir, I. A., Dinata, Y., Linkie, M. & Macdonald, D. W. (2013). Asiatic golden cat and Sunda clouded leopard occupancy in the Kerinci Seblat landscape, West-Central Sumatra. Cat News, 59, 7–10.
Hansen, M. C., Potapov, P. V., Moore, R., Hancher, M., Turubanova, S. A., Tyukavina, A., Thau, D., Stehman, S. V., Goetz, S. J., Loveland, T. R. & Kommareddy, A. (2013). High-Resolution Global Maps of 21st-Century Forest Cover Change. Science. 342(6160), 850–53. https://doi.org/10.1126/science.1244693
Hayward, M. W., Lyngdoh, S. & Habib, B. (2014). Diet and prey preferences of dholes (Cuon alpinus): dietary competition within Asia's apex predator guild. Journal of Zoology, 294(4), 255–266. https://doi.org/10.1111/jzo.12171
Hijmans, R. J., Van Etten, J., Mattiuzzi, M., Sumner, M., Greenberg, J. A., Lamigueiro, O. P., Bevan, A., Racine, E. B. & Shortridge, A. (2013). Raster. R package version 3.6-11.
Jennings, A. P. & Veron, G. (2011). Predicted distributions and ecological niches of civet and mongoose species in Southeast Asia. Journal of Mammalogy, 92(2), 316–327. https://doi.org/10.1644/10-MAMM-A-155.1
Johnsingh, A. J. T. (1992). Prey selection in three sympatric carnivores in Bandipur. Mammalia, 56, 517–526. https://doi.org/10.1515/mamm.1992.56.4.517
Kamler, J. F., Songsasen, N., Jenks, K., Srivathsa, A., Sheng, L. & Kunkel, K. (2015). Cuon alpinus. IUCN Red List of Threatened Species, e.T5953A72477893.
Kawanishi, K. & Sunquist, M. E. (2008). Food habits and activity patterns of the Asiatic golden cat (Catopuma temminckii) and dhole (Cuon alpinus) in a primary rainforest of Peninsular Malaysia. Mammal Study, 33(4), 173–177. https://doi.org/10.3106/1348-6160-33.4.173
Lahkar, D., Ahmed, M. F., Begum, R. H., Das, S. K. & Harihar, A. (2021). Inferring patterns of sympatry among large carnivores in Manas National Park–a prey‐rich habitat influenced by anthropogenic disturbances. Animal Conservation, 24(4), 589–601. https://doi.org/10.1111/acv.12662
Linkie, M., Chapron, G., Martyr, D. J., Holden, J. & Leader-Williams, N. (2006). Assessing the viability of tiger subpopulations in a fragmented landscape. Journal of Applied Ecology, 43(3), 576–586. https://doi.org/10.1111/j.1365-2664.2006.01153.x
MacArthur, R. & Levins, R. (1967). The Limiting Similarity, Convergence, and Divergence of Co-existing Species. The American Naturalist, 101(921), 377–385. https://doi.org/10.1086/282505
Mallegowda, P., Rengaian, G., Krishnan, J. & Niphadkar, M. (2015). Assessing habitat quality of forest-corridors through NDVI analysis in dry tropical forests of south India: implications for conservation. Remote Sensing, 7(2), 1619–1639. https://doi.org/10.3390/rs70201619
Marinho, P. H., Bezerra, D., Antongiovanni, M., Fonseca, C. R. & Venticinque E. M. (2018). Activity patterns of the threatened northern tiger cat Leopardus tigrinus and its potential prey in a Brazilian dry tropical forest. Mammalian Biology, 89, 30–36. https://doi.org/10.1016/j.mambio.2017.12.004
Marneweck, C., Butler, A. R., Gigliotti, L. C., Harris, S. N., Jensen, A. J., Muthersbaugh, M., Newman, B. A., Saldo, E. A., Shute, K., Titus, K. L. & Yu, S. W. (2021). Shining the spotlight on small mammalian carnivores: global status and threats. Biological Conservation, 255, 1009005. https://doi.org/10.1016/j.biocon.2021.109005
May, R., Van Dijk, J., Wabakken, P., Swenson, J. E., Linnell, J. D. C., Zimmermann, B., Odden, J., Pedersen, H. C., Andersen, R. & Landa, A. (2008). Habitat differentiation within the large‐carnivore community of Norway's multiple‐use landscapes. Journal of Applied Ecology, 45(5), 1382–1391. https://doi.org/10.1111/j.1365-2664.2008.01527.x
Menon, V. (2003). A Field Guide to Indian Mammals. Christopher Helm, London, UK, pp.1-200.
Menon, V. (2014). Indian mammals: a field guide. Hachette Book Publishing India Pvt. Ltd., Gurgaon, India, p.528.
Miller, J. R. B. (2010). Survey of western tragopan, koklass pheasant, and Himalayan monal populations in the Great Himalayan National Park, Himachal Pradesh, India. Indian Birds, 6(3), 60–65.
Mohamad, S. W., Rayan, D. M., Christopher, W. C. T., Hamirul, M., Mohamed, A., Lau, C. F. & Siwan, E. S. (2015). The first description of population density and habitat use of the mainland clouded leopard Neofelis nebulosa within a logged-primary forest in South East Asia. Population Ecology, 57(3), 495–503.
https://doi.org/10.1007/s10144-015-0494-1
Nath, A., Singha, H., Haque, M. & Lahkar, B. P. (2019). Sparrows in urban complexity: macro and micro-scale habitat use of sympatric sparrows in Guwahati City, India. Urban Ecosystems, 22(6), 1047–1060.
https://doi.org/10.1007/s11252-019-00876-4
Ngoprasert, D., Lynam, A. J., Sukmasuang, R., Tantipisanuh, N., Chutipong, W., Steinmetz, R., Jenks, K.E., Gale, G. A., Grassman, Jr. L. I., Kitamura, S. & Howard, J. (2012). Occurrence of Three Felids across a Network of Protected Areas in Thailand: Prey, Intraguild, and Habitat Associations. Biotropica, 44(6), 810–817. https://doi.org/10.1111/j.1744-7429.2012.00878.x
O'Brien, T. G., Kinnaird, M. F. & Wibisono, H. T. (2003). Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape. Animal Conservation, 6(2), 131–139. https://doi.org/10.1017/S1367943003003172
Penjor, U., Macdonald, D. W., Wangchuk, S., Tandin, T. & Tan, C. K. W. (2018). Identifying important conservation areas for the clouded leopard Neofelis nebulosa in a mountainous landscape: Inference from spatial modeling techniques. Ecology and Evolution, 8(8), 4278–4291. https://doi.org/10.1002/ece3.3970
Pettorelli, N. (2013). NDVI falls down: exploring situations where it does not work. In: The Normalised Difference Vegetation Index. Oxford Academic, Oxford, UK, 115–126. https://doi.org/10.1093/acprof:osobl/9780199693160.003.0009
Pianka, E. R. (1974). Niche Overlap and Diffuse Competition. Proceedings of the National Academy of Sciences of the United States of America, 71(5), 2141–2145.
https://doi.org/10.1073/pnas.71.5.2141
Prater, S. H. & Barruel, P. (1971). The book of Indian animals. Bombay Natural History Society, Bombay, India, 348.
Rabinowitz, A. R. (1990). Notes on the behaviour and movements of leopard cats, Felis bengalensis, in a dry tropical forest mosaic in Thailand. Biotropica, 22(4), 397–403. https://doi.org/10.2307/2388557
Rabinowitz, A. R. & Walker, R. S. (1991). The carnivore community in a dry tropical forest mosaic in Huai Kha Khaeng Wildlife Sanctuary, Thailand. Journal of Tropical Ecology, 7(1), 37–47. https://doi.org/10.1017/S0266467400005034
Ripple, J., Wirsing, A. J., Wilmers, C. C., Letnic, M (2013). Widespread mesopredator effects after wolf extirpation. Biological Conservation, 160, 70–79.
https://doi.org/10.1016/j.biocon.2012.12.033
Ripple, W. J., Estes, J. A., Beschta, R. L., Wilmers, C. C., Ritchie, E. G., Hebblewhite, M., Berger, J., Elmhagen, B., Letnic, M., Nelson, M. P. & Schmitz, O. J. (2014). Status and Ecological Effects of the World's Largest Carnivores. Science, 343(6167), 1241484–1241484. https://doi.org/10.1126/science.1241484
Rota, C. T., Ferreira, M. A. R., Kays, R. W., Forrester, T. D., Kalies, L., Mcshea, W. J., Parsons, A. W. & Millspaugh, J. J. (2016). A Multispecies Occupancy Model for Two or More Interacting Species. Methods in Ecology and Evolution, 7(10), 1164–1173. https://doi.org/10.1111/2041-210X.12587
Sarma, P. K., Lahkar, B. P., Ghosh, S., Rabha A., Das J. P., Nath, N. K., Dey, S. & Brahma, N. (2008). Land-use and land-cover change and future implication analysis in Manas National Park, India using multi- temporal satellite data. Current Science, 95(2), 223–227.
Sathyakumar, S., Bashir, T., Bhattacharya, T. & Poudyal, K. (2011). Assessing mammal distribution and abundance in intricate eastern Himalayan habitats of Khangchendzonga, Sikkim India. Mammalia, 75(3), 257–268. https://doi.org/10.1515/mamm.2011.023
Sathyakumar, S., Sharma, L. K. & Charoo, S. A. (2013). Ecology of Asiatic Black Bear in Dachigam National Park, Kashmir, India. Final Project Report, Wildlife Institute of India, Dehradun, India.
Selvan, K. M., Veeraswami, G. G. & Hussain, S. A. (2013a). Dietary preference of the Asiatic wild dog (Cuon alpinus). Mammalian Biology, 78(6), 486–489. https://doi.org/10.1016/j.mambio.2013.08.007
Selvan, K. M., Veeraswami, G. G., Lyngdoh, S., Habib, B. & Hussain, S. A. (2013b). Prey selection and food habits of three sympatric large carnivores in a tropical lowland forest of the Eastern Himalayan Biodiversity Hotspot. Mammalian Biology, 78(4), 296–303. https://doi.org/10.1016/j.mambio.2012.11.009
Soberón, J. (2007). Grinnellian and Eltonian niches and geographic distributions of species. Ecology Letters, 10(12), 1115–1123. https://doi.org/10.1111/j.1461-0248.2007.01107.x
Sollmann, R., Mohamed, A., Samejima, H. & Wilting, A. (2013). Risky business or simple solution - Relative abundance indices from camera-trapping. Biological Conservation, 159, 405–412. https://doi.org/10.1016/j.biocon.2012.12.025
Stein, A. B., Athreya, V., Gerngross, P., Balme, G., Henschel, P., Karanth, U., Miquelle, D., Rostro-Garcia, S., Kamler, J. F., Laguardia, A., Khorozyan, I. & Ghoddousi, A. (2020). Panthera pardus (amended version of 2019 assessment). The IUCN Red List of Threatened Species 2020, e. T15954A163991139.
Steinmetz, R., Seuaturien, N. & Chutipong, W. (2013). Tigers, leopards, and dholes in a half-empty forest: assessing species interactions in a guild of threatened carnivores. Biological Conservation, 163, 68–78. https://doi.org/10.1016/j.biocon.2012.12.016
Suraci, J. P., Clinchy, M., Dill, L. M., Roberts, D. & Zanette, L. Y. (2016). Fear of large carnivores causes a trophic cascade. Nature Communications, 7, 10698. https://doi.org/10.1038/ncomms10698
Terborgh, J. & Estes, J. A. (2013). Trophic cascades: predators, prey, and the changing dynamics of nature. Island Press, Washington D.C., USA, pp.1-488.
Thompson, C. M. & Gese, E. M. (2007). Food webs and intraguild predation: community interactions of a native mesocarnivore. Ecology, 88(2), 334–346.
https://doi.org/10.1890/0012-9658(2007)88[334:FWAIPC]2.0.CO;2
Time & Date (2021). Temperature and Humidity. https://timeanddate.com. (20 September 2021).
Tobler, M. W., Carrillo‐Percastegui, S. E., Leite Pitman, R., Mares, R. & Powell, G. (2008). An evaluation of camera traps for inventorying large‐and medium‐sized terrestrial rainforest mammals. Animal conservation 11(3), 169–178. https://doi.org/10.1111/j.1469-1795.2008.00169.x
USGS Earth Explorer (2021). Digital elevation model and normalised difference vegetation index. https://earthexplorer.usgs.gov/. (Accessed on 05 October 2021).
Veech, J. A. (2013). A probabilistic model for analysing species co-occurrence. Global Ecology and Biogeography, 22(2), 252–260. https://doi.org/10.1111/j.1466-8238.2012.00789.x
Vernes, K., Jackson, S. M., Elliott, T. F., Tischler, M. & Harper, A. (2021). Diets of mammalian carnivores in the deserts of north-eastern South Australia. Journal of Arid Environments, 188, 104377. https://doi.org/10.1016/j.jaridenv.2020.104377
Vernes, K., Rajaratnam, R. & Dorji, S. (2022). Patterns of species co‑ occurrence in a diverse Eastern Himalayan montane carnivore community Mammal Research, 67(2), 139–149. https://doi.org/10.1007/s13364-022-00618-6
Wang, S. W. (2001). Conservation management plan for the black mountains national park. Nature Conservation Division. Ministry of Agriculture, Thimphu, Bhutan.
Wasserstein, R. L., Schirm, A. L. & Lazar, N. A. (2019). Moving to a world beyond "p< 0.05". The American Statistician, 73(sup1), 1–19. https://doi.org/10.1080/00031305.2019.1583913
Wibisono, H. T. & McCarthy, J. (2010). Melanistic marbled cat from Bukit Barisan Selatan National Park, Sumatra, Indonesia. Cat News, 52, 9–10.
Wikramanayake, E. D., Dinerstein, E. & Loucks, C. J. (2002). Terrestrial ecoregions of the Indo-Pacific: a conservation assessment, 3 of ecoregions assessments series. Island Press, Washington DC, USA, 643.
Wilson, D. E. & Mittermeier, R. A. (2011). Handbook of the mammals of the world, 2: hoofed mammals. Lynx Ediciones, Barcelona, Spain, pp.1-886.
Zhang, J. L., Ding, Q. & Huang, J. H. (2013). Spaa: Species Association Analysis. R package version 0.2.2.

Edited By
Ajith Kumar
National Centre for Biological Sciences
*CORRESPONDENCE
Salvador Lyngdoh
✉ salvador@wii.gov.in
CITATION
Bhatt, U., Lyngdoh, S. (2024). Carnivore Chronicles: Co-occurrence and habitat use in the tropical forest of Manas National Park, North-East India. Journal of Wildlife Science,1(1), 16-30. https://doi.org/10.63033/JWLS.GCTQ1198
FUNDING
The study was funded by Department of Science and Technology, Govt. of India under grant no. EMR/2015/000085 of 01-04-2016.
COPYRIGHT
© 2024 Bhatt, Lyngdoh. This is an open-access article, immediately and freely available to read, download, and share. The information contained in tarticle is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0![]()
![]() ), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepacademic practice. Copyright is retained by the author(s).
), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepacademic practice. Copyright is retained by the author(s).
PUBLISHED BY
Wildlife Institute of India, Dehradun, 248 001 INDIA
PUBLISHER'S NOTE
The Publisher, Journal of Wildlife Science or Editors cannot be held responsible for any errors or consequences arising from the use of the informatcontained in this article. All claims expressed in this article are solely those of the author(s) and do not necessarily represent those of their affiliated organisations or those of the publisher, the editors and the reviewers. Any prodthat may be evaluated or used in this article or claim made by its manufacturer is not guaranteed or endorsed by the publisher.
Acharya, B. B. (2007). The Ecology of the Dhole or Asiatic Wild Dog (Cuon alpinus) in Pench Tiger Reserve, Madhya Pradesh. PhD dissertation, Saurashtra University, Rajkot, India.
Ancrenaz, M., Hearn, A. J., Ross, J., Sollmann, R. & Wilting, A. (2012). Handbook for Wildlife Monitoring Using Camera-Traps. BBEC Publication, Sabah, Malaysia, 71.
Anderson, D. R. (2008). Model Based Inference in the Life Sciences. Springer, New York, 184. https://doi.org/10.1007/978-0-387-74075-1
Araujo, M. B. & Guisan, A. (2006). Five (or so) challenges for species distribution modelling. Journal of Biogeography, 33(10), 1677–1688. https://doi.org/10.1111/j.1365-2699.2006.01584.x
Ashrafzadeh, M. R., Khosravi, R., Adibi, M. A., Taktehrani, A., Wan, H. Y. & Cushman, S. A. (2020). A multi-scale, multi-species approach for assessing effectiveness of habitat and connectivity conservation for endangered felids. Biological Conservation, 245, 108523. https://doi.org/10.1016/j.biocon.2020.108523
Austin, S. C., Tewes, M. E., Grassman, Jr. L. I. & Silvy, N. J. (2010). Ecology and conservation of the leopard cat Prionailurus bengalensis and clouded leopard Neofelis nebulosa in Khao Yai National Park, Thailand. Acta Zool Sinica, 53, 1–14.
Azlan, J. (2003). The diversity and conservation of mustelids, viverrids, and herpestids in a disturbed forest in Peninsular Malaysia. Small Carnivore Conservation, 29, 8–9.
Barton, K. (2016). MuMIn: Multi-Model Inference. R package version, 1.47.1.
Berentsen, A. R., Rivera-Rodriguez, M. J., McClure, K. M., Torres-Toledo, F. B., Garcia-Cancel, J. G. & Gilbert A. T. (2020). Home range estimates for small Indian mongooses (Urva auropunctata) in southwestern Puerto Rico. Caribbean Journal of Science, 50(2), 225–235. https://doi.org/10.18475/cjos.v50i2.a4
Bhatt, U., Adhikari, B. S., Habib, B. & Lyngdoh, S. (2021). Temporal interactions and moon illumination effect on mammals in a tropical semievergreen forest of Manas National Park, Assam, India. Biotropica, 53(3), 831–845. https://doi.org/10.1111/btp.12917
Bhatt, U., Adhikari, B. S. & Lyngdoh, S. (2022). Monitoring diversity and abundance of mammals with camera-traps: a case study of Manas National Park, Assam, India. Check List, 18(5), 1023–1043. https://doi.org/10.15560/18.5.1023
Bhatt, U. & Lyngdoh, S. (2023a). Secrets of the clouded leopard: abundance, habitat use and carnivore coexistence in tropical forest of Manas National Park, Assam, India. Oryx, 57(6), 757–768. https://doi.org/10.1017/S0030605322001661
Bhatt, U. & Lyngdoh, S. (2023b). Do dholes segregate themselves from their sympatrids? Habitat use and carnivore co-existence in the tropical forest. Mammalian Biology, 103(6), 591–601. https://doi.org/10.1007/s42991-023-00378-z
Bhattacharjya, D. K., Kar, A., Sarma, H. & Patowary, K. N. (2015). Notes on herbal treatment practiced by the people of fringe villages of Manas National Park. India. Indian Journal of Traditional Knowledge, 1(1), 155–160.
Borah, J., Sharma, T., Das, D., Rabha, N., Kakati, N., Basumatary, A., Ahmed, M. F. & Vattakaven, J. (2014). Abundance and density estimates for common leopard Panthera pardus and clouded leopard Neofelis nebulosa in Manas National Park, Assam, India. Oryx, 48(1), 149–155. https://doi.org/10.1017/S0030605312000373
Borah, J., Sharma, T., Das N., Rabha, N., Kakati, N., Basumatri, A., Ahmed, F., Vattakaven, J., Bhobora, C. & Swargowari, A. (2012). Diversity of carnivores in Manas National Park - a World Heritage Site, Assam, India. Cat News, 56, 16–19.
Borah, J., Wangchuk, D., Swargowari, A., Wangchuk, T., Sharma, T., Das, D., Rabha, N., Basumatari A., Kakati N. et al. (2013). Tigers in the Transboundary Manas Conservation Complex: conservation implications across borders. Parks, 19, 51–62.
Brodie, J. & Giordano, A. J. (2012). Density of the Vulnerable Sunda clouded leopard Neofelis diardi in a protected area in Sabah, Malaysian Borneo. Oryx, 46(3), 427–430. https://doi.org/10.1017/S0030605312000087
Carbone, C., Christie, S., Conforti, K., Coulson, T., Franklin, N., Ginsberg, J. R., Griffiths, M., Holden, J., Kawanishi, K. et al. (2001) The use of photographic rates to estimate densities of tigers and other cryptic mammals. Animal Conservation, 4 (1), 75–79. https://doi.org/10.1017/S1367943001001081
Chatterjee, N., Nigam, P. & Habib, B. (2020). Population density and habitat use of two sympatric small cats in a central Indian reserve. Plos One, 15(6), e0233569. https://doi.org/10.1371/journal.pone.0233569
Chaudhary, R., Zehra, N., Musavi, A. & Khan, J. A. (2020). Evaluating the effect of ecological and anthropogenic variables on site use by sympatric large carnivores in Gir protected area, Gujarat, India. Wildlife Biology, 4, 1–7. https://doi.org/10.2981/wlb.00696
Choudhury, A. (2013). The mammals of North east India. Gibbon Books and the Rhino Foundation for Nature in NE India, Guwahati, Assam, India, 432.
Choudhury, A., Wozencraft, C., Muddapa, D., Yonzon, P., Jennings, A. & Geraldine, V. (2011). Herpestes edwardsii. In: IUCN 2012. IUCN Red List of Threatened Species, version 2012.2. www.iucnredlist.org. (Accessed on 03 November 2012).
Chutipong, W., Steinmetz, R., Savini, T. & Gale, G. A. (2017). Assessing resource and predator effects on habitat use of tropical small carnivores. Mammal Research, 62(1), 21–36. https://doi.org/10.1007/s13364-016-0283-z
Chutipong, W., Tantipisanuh, N., Ngoprasert, D., Lynam, A. J., Steinmetz, R., Jenks, K. E., Grassman, Jr. L. I, Tewes, M., Kitamura, S. & Baker, M. C. (2014). Current distribution and conservation status of small carnivores in Thailand: a baseline review. Small Carnivore Conservation, 51, 96–136.
Corbet, G. B. & Hill, J. E. (1992). The mammals of the Indo-Malayan region: a systematic review. Oxford University Press, Oxford, and Natural History Museum, London, 496.
Davis, M. L., Kelly, M. J. & Stauffer, D. F. (2011). Carnivore co-existence and habitat use in the Mountain Pine Ridge Forest Reserve, Belize. Animal Conservation, 14(1), 56–65. https://doi.org/10.1111/j.1469-1795.2010.00389.x
DebRoy, S. (1991). Manas: a monograph. Tiger paper (FAO), 18, 6–15.
Di Bitetti, M. S., De Angelo, C. D., Di Blanco, Y. E. & Paviolo, A. (2010). Niche partitioning and species co-existence in a Neotropical felid assemblage. Acta Oecologica, 36(4), 403– 412. https://doi.org/10.1016/j.actao.2010.04.001
Duckworth, J. W., Timmins, R. J., Choudhury, A., Chutipong, W., Willcox, D. H. A., Mudappa, D., Rahman H., Widmann, P., Wilting, A. & Xu, W. (2016). Paradoxurus hermaphroditus (Pallas, 1777). The IUCN Red List of Threatened Species 2016.
Elfström, M., Zedrosser, A., Støen, O. G. & Swenson, J. E. (2014). Ultimate and proximate mechanisms underlying the occurrence of bears close to human settlements: review and management implications. Mammal Review, 44(1), 5–18. https://doi.org/10.1111/j.1365-2907.2012.00223.x
ESRI (2011). ArcGIS Desktop: Release 10. Environmental System Research Institute, Inc., Redlands, California, USA.
Garshelis D. & Steinmetz R. (2020) Ursus thibetanus (amended version of 2016 assessment). The IUCN Red List of Threatened Species 2020: e.T22824A166528664.
George, S. J. (1994). The Bodo movement in Assam: Unrest to accord. Asian Survey, 34(10), 878–892. https://doi.org/10.2307/2644967
Glen, A. S. & Dickman, C. R. (2005). Complex interactions among mammalian carnivores in Australia, and their implications for wildlife management. Biological Reviews, 80(3), 387–401. https://doi.org/10.1017/S1464793105006718
Goldman, H. V. & Winther-Hansen, J. (2003). First photographs of the Zanzibar servaline Genet Genetta servalina archeri and other endemic subspecies on the island of Unguja, Tanzania. Small Carnivore Conservation, 29, 1–4.
Gompper, M. E., Lesmeister, D. B., Ray, J. C., Malcolm, J. R. & Kays, R. (2016). Differential Habitat Use or Intraguild Interactions: What Structures a Carnivore Community? Plos One, 11(1), e0146055. https://doi.org/10.1371/journal.pone.0146055
Goodrich, J., Wibisono, H., Miquelle, D., Lynam, A. J., Sanderson, E., Chapman, S., Gray, T. N. E., Chanchani, P. & Harihar, A. (2022). Panthera tigris. The IUCN Red List of Threatened Species, 2022, e.T15955A214862019.
Goswami R. & Ganesh, T. (2014). Carnivore and herbivore densities in the immediate aftermath of ethno-political conflict: the case of Manas National Park, India. Tropical Conservation Science, 7(3), 475–487. https://doi.org/10.1177/194008291400700308
Grassman, Jr. L. I, Tewes, M. E., Silvy, N. J. & Kreetiyutanont, K. (2005). Ecology of three sympatric felids in a mixed evergreen forest in north-central Thailand. Journal of Mammalogy, 86, 29–38. https://doi.org/10.1644/1545-1542(2005)086<0029:EOTSFI>2.0.CO;2
Griffith, D. M., Veech, J. A. & Marsh, C. J. (2016). Co-occur: Probabilistic species co-occurrence analysis in R. Journal of Statistical Software, 69(2), 1–17. https://doi.org/10.18637/jss.v069.c02
Griffiths, M. & Van Schaick, C. P. (1993). Camera-trapping: A new tool for the study of elusive rain forest animals. Tropical Biodiversity, 1, 131–135.
Haidir, I. A., Dinata, Y., Linkie, M. & Macdonald, D. W. (2013). Asiatic golden cat and Sunda clouded leopard occupancy in the Kerinci Seblat landscape, West-Central Sumatra. Cat News, 59, 7–10.
Hansen, M. C., Potapov, P. V., Moore, R., Hancher, M., Turubanova, S. A., Tyukavina, A., Thau, D., Stehman, S. V., Goetz, S. J., Loveland, T. R. & Kommareddy, A. (2013). High-Resolution Global Maps of 21st-Century Forest Cover Change. Science. 342(6160), 850–53. https://doi.org/10.1126/science.1244693
Hayward, M. W., Lyngdoh, S. & Habib, B. (2014). Diet and prey preferences of dholes (Cuon alpinus): dietary competition within Asia's apex predator guild. Journal of Zoology, 294(4), 255–266. https://doi.org/10.1111/jzo.12171
Hijmans, R. J., Van Etten, J., Mattiuzzi, M., Sumner, M., Greenberg, J. A., Lamigueiro, O. P., Bevan, A., Racine, E. B. & Shortridge, A. (2013). Raster. R package version 3.6-11.
Jennings, A. P. & Veron, G. (2011). Predicted distributions and ecological niches of civet and mongoose species in Southeast Asia. Journal of Mammalogy, 92(2), 316–327. https://doi.org/10.1644/10-MAMM-A-155.1
Johnsingh, A. J. T. (1992). Prey selection in three sympatric carnivores in Bandipur. Mammalia, 56, 517–526. https://doi.org/10.1515/mamm.1992.56.4.517
Kamler, J. F., Songsasen, N., Jenks, K., Srivathsa, A., Sheng, L. & Kunkel, K. (2015). Cuon alpinus. IUCN Red List of Threatened Species, e.T5953A72477893.
Kawanishi, K. & Sunquist, M. E. (2008). Food habits and activity patterns of the Asiatic golden cat (Catopuma temminckii) and dhole (Cuon alpinus) in a primary rainforest of Peninsular Malaysia. Mammal Study, 33(4), 173–177. https://doi.org/10.3106/1348-6160-33.4.173
Lahkar, D., Ahmed, M. F., Begum, R. H., Das, S. K. & Harihar, A. (2021). Inferring patterns of sympatry among large carnivores in Manas National Park–a prey‐rich habitat influenced by anthropogenic disturbances. Animal Conservation, 24(4), 589–601. https://doi.org/10.1111/acv.12662
Linkie, M., Chapron, G., Martyr, D. J., Holden, J. & Leader-Williams, N. (2006). Assessing the viability of tiger subpopulations in a fragmented landscape. Journal of Applied Ecology, 43(3), 576–586. https://doi.org/10.1111/j.1365-2664.2006.01153.x
MacArthur, R. & Levins, R. (1967). The Limiting Similarity, Convergence, and Divergence of Co-existing Species. The American Naturalist, 101(921), 377–385. https://doi.org/10.1086/282505
Mallegowda, P., Rengaian, G., Krishnan, J. & Niphadkar, M. (2015). Assessing habitat quality of forest-corridors through NDVI analysis in dry tropical forests of south India: implications for conservation. Remote Sensing, 7(2), 1619–1639. https://doi.org/10.3390/rs70201619
Marinho, P. H., Bezerra, D., Antongiovanni, M., Fonseca, C. R. & Venticinque E. M. (2018). Activity patterns of the threatened northern tiger cat Leopardus tigrinus and its potential prey in a Brazilian dry tropical forest. Mammalian Biology, 89, 30–36. https://doi.org/10.1016/j.mambio.2017.12.004
Marneweck, C., Butler, A. R., Gigliotti, L. C., Harris, S. N., Jensen, A. J., Muthersbaugh, M., Newman, B. A., Saldo, E. A., Shute, K., Titus, K. L. & Yu, S. W. (2021). Shining the spotlight on small mammalian carnivores: global status and threats. Biological Conservation, 255, 1009005. https://doi.org/10.1016/j.biocon.2021.109005
May, R., Van Dijk, J., Wabakken, P., Swenson, J. E., Linnell, J. D. C., Zimmermann, B., Odden, J., Pedersen, H. C., Andersen, R. & Landa, A. (2008). Habitat differentiation within the large‐carnivore community of Norway's multiple‐use landscapes. Journal of Applied Ecology, 45(5), 1382–1391. https://doi.org/10.1111/j.1365-2664.2008.01527.x
Menon, V. (2003). A Field Guide to Indian Mammals. Christopher Helm, London, UK, pp.1-200.
Menon, V. (2014). Indian mammals: a field guide. Hachette Book Publishing India Pvt. Ltd., Gurgaon, India, p.528.
Miller, J. R. B. (2010). Survey of western tragopan, koklass pheasant, and Himalayan monal populations in the Great Himalayan National Park, Himachal Pradesh, India. Indian Birds, 6(3), 60–65.
Mohamad, S. W., Rayan, D. M., Christopher, W. C. T., Hamirul, M., Mohamed, A., Lau, C. F. & Siwan, E. S. (2015). The first description of population density and habitat use of the mainland clouded leopard Neofelis nebulosa within a logged-primary forest in South East Asia. Population Ecology, 57(3), 495–503.
https://doi.org/10.1007/s10144-015-0494-1
Nath, A., Singha, H., Haque, M. & Lahkar, B. P. (2019). Sparrows in urban complexity: macro and micro-scale habitat use of sympatric sparrows in Guwahati City, India. Urban Ecosystems, 22(6), 1047–1060.
https://doi.org/10.1007/s11252-019-00876-4
Ngoprasert, D., Lynam, A. J., Sukmasuang, R., Tantipisanuh, N., Chutipong, W., Steinmetz, R., Jenks, K.E., Gale, G. A., Grassman, Jr. L. I., Kitamura, S. & Howard, J. (2012). Occurrence of Three Felids across a Network of Protected Areas in Thailand: Prey, Intraguild, and Habitat Associations. Biotropica, 44(6), 810–817. https://doi.org/10.1111/j.1744-7429.2012.00878.x
O'Brien, T. G., Kinnaird, M. F. & Wibisono, H. T. (2003). Crouching tigers, hidden prey: Sumatran tiger and prey populations in a tropical forest landscape. Animal Conservation, 6(2), 131–139. https://doi.org/10.1017/S1367943003003172
Penjor, U., Macdonald, D. W., Wangchuk, S., Tandin, T. & Tan, C. K. W. (2018). Identifying important conservation areas for the clouded leopard Neofelis nebulosa in a mountainous landscape: Inference from spatial modeling techniques. Ecology and Evolution, 8(8), 4278–4291. https://doi.org/10.1002/ece3.3970
Pettorelli, N. (2013). NDVI falls down: exploring situations where it does not work. In: The Normalised Difference Vegetation Index. Oxford Academic, Oxford, UK, 115–126. https://doi.org/10.1093/acprof:osobl/9780199693160.003.0009
Pianka, E. R. (1974). Niche Overlap and Diffuse Competition. Proceedings of the National Academy of Sciences of the United States of America, 71(5), 2141–2145.
https://doi.org/10.1073/pnas.71.5.2141
Prater, S. H. & Barruel, P. (1971). The book of Indian animals. Bombay Natural History Society, Bombay, India, 348.
Rabinowitz, A. R. (1990). Notes on the behaviour and movements of leopard cats, Felis bengalensis, in a dry tropical forest mosaic in Thailand. Biotropica, 22(4), 397–403. https://doi.org/10.2307/2388557
Rabinowitz, A. R. & Walker, R. S. (1991). The carnivore community in a dry tropical forest mosaic in Huai Kha Khaeng Wildlife Sanctuary, Thailand. Journal of Tropical Ecology, 7(1), 37–47. https://doi.org/10.1017/S0266467400005034
Ripple, J., Wirsing, A. J., Wilmers, C. C., Letnic, M (2013). Widespread mesopredator effects after wolf extirpation. Biological Conservation, 160, 70–79.
https://doi.org/10.1016/j.biocon.2012.12.033
Ripple, W. J., Estes, J. A., Beschta, R. L., Wilmers, C. C., Ritchie, E. G., Hebblewhite, M., Berger, J., Elmhagen, B., Letnic, M., Nelson, M. P. & Schmitz, O. J. (2014). Status and Ecological Effects of the World's Largest Carnivores. Science, 343(6167), 1241484–1241484. https://doi.org/10.1126/science.1241484
Rota, C. T., Ferreira, M. A. R., Kays, R. W., Forrester, T. D., Kalies, L., Mcshea, W. J., Parsons, A. W. & Millspaugh, J. J. (2016). A Multispecies Occupancy Model for Two or More Interacting Species. Methods in Ecology and Evolution, 7(10), 1164–1173. https://doi.org/10.1111/2041-210X.12587
Sarma, P. K., Lahkar, B. P., Ghosh, S., Rabha A., Das J. P., Nath, N. K., Dey, S. & Brahma, N. (2008). Land-use and land-cover change and future implication analysis in Manas National Park, India using multi- temporal satellite data. Current Science, 95(2), 223–227.
Sathyakumar, S., Bashir, T., Bhattacharya, T. & Poudyal, K. (2011). Assessing mammal distribution and abundance in intricate eastern Himalayan habitats of Khangchendzonga, Sikkim India. Mammalia, 75(3), 257–268. https://doi.org/10.1515/mamm.2011.023
Sathyakumar, S., Sharma, L. K. & Charoo, S. A. (2013). Ecology of Asiatic Black Bear in Dachigam National Park, Kashmir, India. Final Project Report, Wildlife Institute of India, Dehradun, India.
Selvan, K. M., Veeraswami, G. G. & Hussain, S. A. (2013a). Dietary preference of the Asiatic wild dog (Cuon alpinus). Mammalian Biology, 78(6), 486–489. https://doi.org/10.1016/j.mambio.2013.08.007
Selvan, K. M., Veeraswami, G. G., Lyngdoh, S., Habib, B. & Hussain, S. A. (2013b). Prey selection and food habits of three sympatric large carnivores in a tropical lowland forest of the Eastern Himalayan Biodiversity Hotspot. Mammalian Biology, 78(4), 296–303. https://doi.org/10.1016/j.mambio.2012.11.009
Soberón, J. (2007). Grinnellian and Eltonian niches and geographic distributions of species. Ecology Letters, 10(12), 1115–1123. https://doi.org/10.1111/j.1461-0248.2007.01107.x
Sollmann, R., Mohamed, A., Samejima, H. & Wilting, A. (2013). Risky business or simple solution - Relative abundance indices from camera-trapping. Biological Conservation, 159, 405–412. https://doi.org/10.1016/j.biocon.2012.12.025
Stein, A. B., Athreya, V., Gerngross, P., Balme, G., Henschel, P., Karanth, U., Miquelle, D., Rostro-Garcia, S., Kamler, J. F., Laguardia, A., Khorozyan, I. & Ghoddousi, A. (2020). Panthera pardus (amended version of 2019 assessment). The IUCN Red List of Threatened Species 2020, e. T15954A163991139.
Steinmetz, R., Seuaturien, N. & Chutipong, W. (2013). Tigers, leopards, and dholes in a half-empty forest: assessing species interactions in a guild of threatened carnivores. Biological Conservation, 163, 68–78. https://doi.org/10.1016/j.biocon.2012.12.016
Suraci, J. P., Clinchy, M., Dill, L. M., Roberts, D. & Zanette, L. Y. (2016). Fear of large carnivores causes a trophic cascade. Nature Communications, 7, 10698. https://doi.org/10.1038/ncomms10698
Terborgh, J. & Estes, J. A. (2013). Trophic cascades: predators, prey, and the changing dynamics of nature. Island Press, Washington D.C., USA, pp.1-488.
Thompson, C. M. & Gese, E. M. (2007). Food webs and intraguild predation: community interactions of a native mesocarnivore. Ecology, 88(2), 334–346.
https://doi.org/10.1890/0012-9658(2007)88[334:FWAIPC]2.0.CO;2
Time & Date (2021). Temperature and Humidity. https://timeanddate.com. (20 September 2021).
Tobler, M. W., Carrillo‐Percastegui, S. E., Leite Pitman, R., Mares, R. & Powell, G. (2008). An evaluation of camera traps for inventorying large‐and medium‐sized terrestrial rainforest mammals. Animal conservation 11(3), 169–178. https://doi.org/10.1111/j.1469-1795.2008.00169.x
USGS Earth Explorer (2021). Digital elevation model and normalised difference vegetation index. https://earthexplorer.usgs.gov/. (Accessed on 05 October 2021).
Veech, J. A. (2013). A probabilistic model for analysing species co-occurrence. Global Ecology and Biogeography, 22(2), 252–260. https://doi.org/10.1111/j.1466-8238.2012.00789.x
Vernes, K., Jackson, S. M., Elliott, T. F., Tischler, M. & Harper, A. (2021). Diets of mammalian carnivores in the deserts of north-eastern South Australia. Journal of Arid Environments, 188, 104377. https://doi.org/10.1016/j.jaridenv.2020.104377
Vernes, K., Rajaratnam, R. & Dorji, S. (2022). Patterns of species co‑ occurrence in a diverse Eastern Himalayan montane carnivore community Mammal Research, 67(2), 139–149. https://doi.org/10.1007/s13364-022-00618-6
Wang, S. W. (2001). Conservation management plan for the black mountains national park. Nature Conservation Division. Ministry of Agriculture, Thimphu, Bhutan.
Wasserstein, R. L., Schirm, A. L. & Lazar, N. A. (2019). Moving to a world beyond "p< 0.05". The American Statistician, 73(sup1), 1–19. https://doi.org/10.1080/00031305.2019.1583913
Wibisono, H. T. & McCarthy, J. (2010). Melanistic marbled cat from Bukit Barisan Selatan National Park, Sumatra, Indonesia. Cat News, 52, 9–10.
Wikramanayake, E. D., Dinerstein, E. & Loucks, C. J. (2002). Terrestrial ecoregions of the Indo-Pacific: a conservation assessment, 3 of ecoregions assessments series. Island Press, Washington DC, USA, 643.
Wilson, D. E. & Mittermeier, R. A. (2011). Handbook of the mammals of the world, 2: hoofed mammals. Lynx Ediciones, Barcelona, Spain, pp.1-886.
Zhang, J. L., Ding, Q. & Huang, J. H. (2013). Spaa: Species Association Analysis. R package version 0.2.2.