TYPE: Research Article ![]()
Avadhoot D. Velankar¹,², Honnavalli N. Kumara¹, V.R. Sheethal¹, Arijit Pal¹,³, Partha Sarathi Mishra¹,⁴, Mewa Singh⁵*
¹Salim Ali Centre for Ornithology and Natural History, South India Centre of Wildlife Institute of India, Anaikatty Post, Coimbatore 641108, Tamil Nadu, India
²Wildlife Conservation Society India, 551, 7th Main Road Rajiv Gandhi Nagar, 2nd Phase Bengaluru - 560097 Karnataka, India
³The Food and Land Use Coalition India, WRI India, AADI, Hauz Khas, New Delhi 110016, India
⁴Srishti Manipal Institute of Arts, Design and Technology, MAHE Bengaluru Campus, Govindapura, Bengaluru–560064, Karnataka, India
⁵Biopsychology Laboratory, Institution of Excellence, Vijnana Bhavan, University of Mysore, Manasagangotry, Mysore-570006, Karnataka, India

Introduction
Two aspects of an organism’s behavioural profile that are essential for its overall adaptation are the way an organism allots its time to various activities and the pattern of its habitat use. Unlike resources such as food, mate, and space, time is constant and finite throughout the lifecycle of the organism. Animals budget their time and energy for different activities in an adaptive manner. Wilson (1975) made two generalizations about time-activity-energy budgets: the principle of stringency stating that these budgets evolve to fit the times of greatest stringency, and the principle of allocation that time and energy are allocated to activities in terms of priorities that usually descend as food, antipredation, and reproduction. Monitoring the activity budget of an animal is the easiest way to understand the interdependence of the animal and its environment. Hence, spatio-temporal changes in environment, physiology, and social organization of a species are directly reflected in changes in time invested in various activities. For example, barbary macaques (Macaca sylvanus) spent more time feeding during snowfall days than during no snowfall (Majolo et. al., 2013). Urban commensal rhesus macaque (M. mulatta) groups spent more time resting than rural groups (Jaman & Huffman, 2013). Similarly, the diet pattern of a species dictates its activity budget. A frugivorous and habitat specialist species such as the lion-tailed macaque (M. silenus) was found to move farther in search of food during the resource crunch period than during the resource-rich period (Santosh et. al., 2015). On the other hand, folivorous species such as Guereza (Colobus guereza) and the snub-nosed monkey (Rhinopithecus roxellana) were found to spend more time resting than moving (Fashing, 2001; Guo et. al., 2007). The social organization of a species also influences the time spent on different activities. Species that are involved in scramble competition over limited food resources tend to spend more time moving (van Schaik et. al., 1983) than species without scramble competition (Fashing, 2001).
Changes in diurnal activity due to environmental, physiological, or social factors manifest as variations in ranging patterns within and between species. Lion-tailed macaques were reported to use regions with high fruit availability and shifted their home range accordingly during the wet and dry seasons (Santosh et. al., 2015). Similar results were reported in red colobus (Colobus badius), where time spent in the area correlated with availability of food resources (Clutton-Brock, 1975). Habitat use is also observed to vary seasonally. Seasonal shrinkage in home range size was observed in Japanese macaques (M. fuscata) due to the availability of supplemental food during the tourist season (Koganezawa & Imaki, 1999). In lion-tailed macaques, ranging varied seasonally, which was influenced by the availability of Artocarpus heterophyllus and Ficus amplissima in summer, Artocarpus heterophyllus in monsoon, and Cullenia exarillata and Toona ciliata in post-monsoon (Erinejery et. al., 2015). Just like the within-group scramble competition affects ranging patterns (van Schaik et. al., 1983), the between-group competition has also been observed to play an important role. Nilgiri langurs (Semnopithecus johnii) were observed to shift their home range to avoid between-group encounters (Kavana et. al., 2014). The availability of food also influences tolerance between neighbouring groups. Japanese macaque was reported to have more overlapping home ranges in resource-rich areas than in resource-poor areas (Maruhashi et. al., 1998).
The long-tailed macaque (M. fascicularis) is a primarily frugivorous species that exploits various resources indicating its generalist nature (Yeager, 1996). They exhibit scramble competition and tend to have larger home ranges for larger group sizes (van Schaik et. al., 1983). The Nicobar Islands in India harbour a unique subspecies of long-tailed macaques (M. f. umbrosus). The 2004 tsunami destroyed large parts of the habitat of these macaques and drastically reduced their population (Sivakumar, 2010). However, over the next decade or so, the population recovered with a very high rate of turnover (Velankar et. al., 2016). Whereas several studies on demography (Pal et. al., 2018a), foraging (Pal et. al., 2018b; Das et. al., 2020; Venlankar et. al., 2023) and social behaviour (Pal et. al., 2019; Mishra et. al., 2020a,b) of this island population arereported, there is no study on their basic activity patterns and habitat use. Here, we report the activity budget and habitat use by these long-tailed macaques inhabiting an area populated by humans at Campbell Bay in the Nicobar Islands and discuss the effect of such habitat on the activity patterns.
Methods
Study Site:
The study was conducted at Campbell Bay, Great Nicobar Island, lying between 7° 1′ 30″ – 7° 0′ 0″ N and 93° 4′ 30″ – 93° 57′ 0″ E (Figure 1). The study site includes a mosaic of natural as well as human-altered land cover types. The coastal region of the study area is dominated by Pandanus spp. and a littoral forest consisting of Ficus rumphii, Alstonia spp., Garcinia nervosa, Terminalia spp., and Semecarpus spp., while the inland region is dominated by human habitation and plantations of coconut (Cocos nucifera) and beetle nut (Areca catechu) (Sinha et. al., 1999). Human habitation consists primarily of small single-story households with kitchen gardens intermittent with open and dense secondary scrub vegetation. The area inhabited by humans is dominated by introduced vegetation consisting of bananas (Musa paradisiaca), guava (Psidium guajava), and mangoes (Mangifera indica). The altitude of the study area ranges from 0 m asl to 218 m asl. Study area receives an annual rainfall of about 3200 mm. A major portion of the rainfall is received from May to November, whereas December to April remains relatively dry.
Study Group:
We selected a group of 22 individuals of long-tailed macaques located at Campbell Bay for the current study. The group had three adult males, seven adult females, two subadult males, seven juveniles, and three infants. The group was habituated for observer presence from April 2013 to October 2013.
Data collection:
The group was followed from dawn (0600 hrs) to dusk (1800 hrs). We recorded the major activities using instantaneous scan sampling (Altmann, 1974) for five minutes at intervals of 30 minutes as per the ethogram provided in Table 1. All individuals in the group were scanned. Though the number of scans for the adults was about equal, it varied among younger individuals due to the age differences and birth intervals. The average number of scans per individual was 5.11 ± 1.9SD. In addition to the major activities, we recorded data on the strata used, feeding, and proximity to the nearest and farthest individuals of the focal animal. For feeding activity during continuous feeding bouts as well as during foraging, details of plant species along with parts consumed and processing and handling by a macaque were documented using Ad-libitum notes. We recorded the geo-coordinates of the visually approximated group centre using a handheld GPS receiver (Garmin GPS Map 76csx) every 30 min.
Analysis
Activity budgets:
We sorted the scan data season-wise, and prepared contingency tables as activity vs month and time of the day for dry and wet seasons. The proportion of each activity was calculated using the aggregated activity budget method (Marsh, 1981). Similarly, information on feeding habits was prepared in contingency tables for the wet and dry seasons. We classified the food items based on the source as plantation and forest species. Food items were further divided into origins of the source i.e. animal, plant, and human waste. The resources of the plant species were subdivided into various parts, such as fruit, leaf, stem, flower, and roots. The proportion of major behavioural states (Table 1) was calculated for a day by dividing the day into 6 classes. We performed the Kruskal-Wallis nonparametric test (Quinn & Keough, 2002) to compare the proportions of the various activities between seasons. We employed the G-test to check for differences within behavioural and feeding classes, and the Z-test of proportions for the difference between seasons.
Habitat Use:
We converted the group locations as shape files. The study area was overlaid with a vector grid of 100 m x 100 m cell dimensions (Grueter et. al., 2009; Santosh et. al., 2015). We kept a one-

Figure 1: Map of the study site in Campbell Bay region with home range of the study group
Table 1: Ethogram of the major behavioural states of the Nicobar long-tailed macaques recorded during the study period along with a brief description03Velankar

hectare area as an appropriate unit to understand the habitat used by the group, considering the home ranges of a few groups (see Pal et. al., 2018). The frequency of grid cell use was computed by counting the number of point locations within a grid cell for wet and dry seasons. The frequencies were converted into proportions by dividing the count of points within a grid cell by the total number of points recorded during each season. The study area was classified into six broad land cover types as habitats viz., Beach, Household, Orchards, Primary Forest, Open scrub, and Plantation. Since the study area was small, land-use and landcover classes were mapped accurately by digitizing them using high-resolution google earth imagery (Figure 1). Ground control points to determine boundarieswere taken using handheld GPS. The spatial availability of each habitat type was estimated as a proportion of habitat type within the area of each grid cell. While following the study group, we recorded the geocoordinate of the group location every 30 minutes. Connecting those consecutive points of a day with the nearest distance was considered the daily path length.
Variation in intensity of grid use and variation in day activity budget between seasons was compared using non-parametric Wilcoxon’s Signed Rank test. We employed McNemar’s chi-square test (Quinn & Keough, 2002) to determine whether the group used the same grids during different seasons. We employed one-way ANOVA to assess variations in the spatial availability of habitat types within each grid cell. The comparisons between classes were done by Tukey’s post-hoc test of Honest Significant Difference (HSD). We used Pearson’s correlation to check for the association between the intensity of grid use and the proportion of available habitat type. Tree density and food species density were also calculated for each grid. Proximity from human settlements, home range overlap with other groups, and the number of between-group encounters observed in each grid were taken as explanatory variables. We used a Generalised Linear Model to test the influence of these seven covariates on the grid-use pattern by the macaques. All statistical analyses were carried out using R statistical language v3.02 (R Development Core Team, 2008) with R Studio IDE for R v0.98.953 with a p-value set at 0.05. We used QGIS v2.13, to prepare land use and land cover maps, as well as to extract information on ranging.
Results
Activity budgets: We collected a total of 4628 individual scans from October 2013 to December 2015 (Table 2). Macaques spent more time resting, which was followed by feeding/foraging, social, and movement, while the time spent on scouting was much shorter (Figure 2) (Kruskal – Wallis χ2 = 53.84, df = 4, P < 0.001). Time spent on each activity was almost similar at different times of the day during the dry (Figure 2a) and the wet (Figure 2b) seasons as indicated by the flatness of the curves. There were no significant difference in activities within each daytime class between seasons (Table 2) (Movement: W = 17, P = 0.17, Feed/forage: W = 10, P = 0.50, , Social: W = 14, P = 0.07, Rest: W = 12, P = 0.75, Scout: W = 17, P = 1.70) (Table2) (Figure5).
Feeding/foraging: The macaques were observed to forage and feed on 26 major food items, of which 12 were introduced by humans and 14 were of native origin (Table 3). The relative frequency of feeding on introduced species was significantly higher (Z = 27.93, P < 0.01) than on native species (Figure 3). Macaques relied significantly more on plant resources than on animal or human waste resources (Z = 19.78, P < 0.01) (Figure 4). Among plant resources, macaques fed on fruits (N = 675) significantly more than flowers (N = 6), stems (N = 32) and leaves (N = 50) (Figure 5) (Z = 15.38, P < 0.01). Coconut (27.0-27.0%), banana (11.9-9.3%), guava (18.6-19.0%), and Pandanus spp.(10.2-8.6%) were the most commonly consumed fruits.
Habitat use: A total of 803 group locations (Dry N = 330, Wet N = 473) were spread over 66 grid cells. Although the total home range size was 66 ha (66 grid cells), the group used only about 6 ha (six grid cells; 9.09%) for most of their movement (Figure 6a). The number of grid cells entered in the dry season (Figure 6b) was more than in the wet season (McNemar’s χ2 = 16.36, p <.01) (Figure 6c). However, the number of intensively used grid cells between the two seasons was almost the same. The group did not use many grids in the western, northern, and southern parts during the wet season. The daily path length was longer in the wet season than in the dry season (t = 2.35, df = 55, p < 0.05) (Table 4).
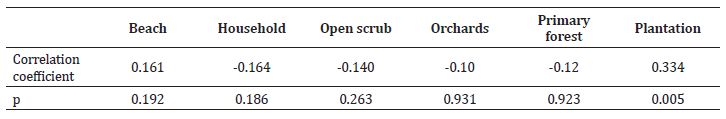
The GLM was run with seven predictors keeping the grid use as the response variable. The model was significantly explanatory when compared with a null model (residual deviance = 913.19, null deviance = 1642.58, DF = 65, residual DF = 61, p < 0.001). The model showed the affinity of the group to grids with overlapping home ranges of adjacent groups. The group also did not avoid the area with frequent between-group encounters (Table 5).
The spatial availability of all habitat types within each grid cell differed significantly (F5, 390 = 19.11, P < 0.001). The mean spatial availability of Primary Forest (0.39 ± 0.40) and Orchards (0.32 ± 0.38) was significantly higher than that of other habitat types (Figure 7). Overall proportion of each habitat type within the home range was 2.90% for beach, 7.70% for households, 14.00% for open scrub, 33.70% for orchards, 38.50% for primary forest and 3.10% for plantation. However, Pearson’s correlation between habitats and the proportion of grid use did not show significant association between all habitats and the intensity of use, except for the plantation which showed weak but significant association (Table 6).
Discussion
The study group spent most of its time on resting which was followed by feeding/foraging, social, and movement. Diurnal activity patterns did not vary among different time slots of the day and between seasons. Although the macaque diet included several food components, its primary diet was fruits. No seasonal variation in dietary patterns was observed. The home range of the study group was 66 ha, while only 6 ha of that was used most frequently. The area used in the wet season was less than the area used in the dry season. The group showed no tendency to avoid overlapping ranges with other groups or areas of greater between-group encounters.
The study group, being partially commensal, largely fed on food resources from orchards and plantations, spending little time in the acquisition of low-cost high-energy foods with ample spare time for resting. Similar observations were reported in rhesus macaque (M. mulatta), where an urban commensal group spent 46% of its time resting alone (Jaman & Huffman, 2013). Similarly, provisioned groups of Macaca fascicularis in Vietnam were observed to spend 31.60% to 36.83% of time resting, which again was the most common activity (Son, 2003). Likewise, a provisioned group of Tibetan macaques (M. thibetana) spent 59.72% of its time resting (Wang et. al., 2007). The time spent on different activities remained almost constant throughout the day. Unlike many other primate species, especially in their natural habitats, which generally show two foraging peaks during a day (Qihai et. al., 2014; Zhou et. al., 2022) and seasonal variations in their time-activity budgets (Zhou et. al., 2007, 2022), the present study group showed no such patterns due to the constant year-round food supply in the human-dominated habitat.
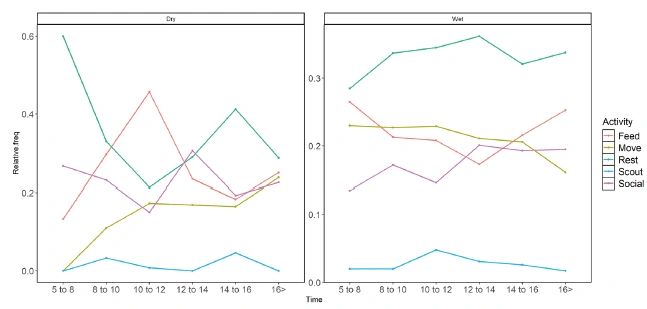
Figure 2. Activities at different times of the day. Left – Dry Season, Right- Wet Season
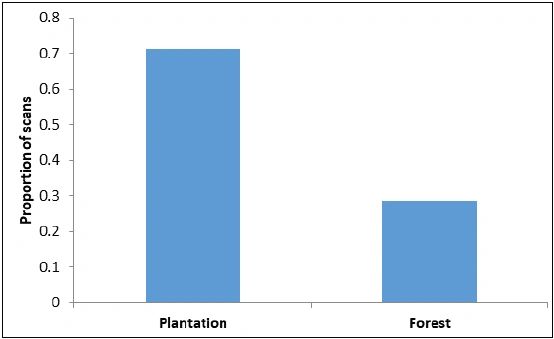
Figure 3: Proportion of food items consumed by Nicobar long tailed macaques
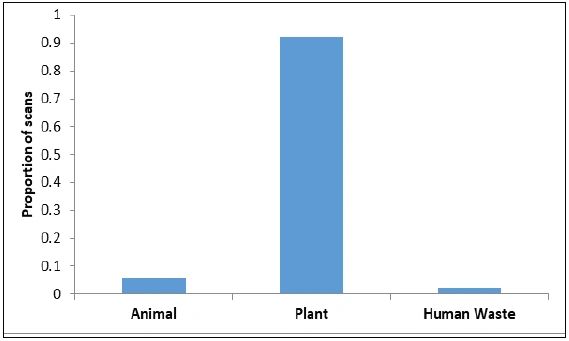
Figure 4: Proportion of food items of animal, plant or human origin consumed by Nicobar long tailed macaques
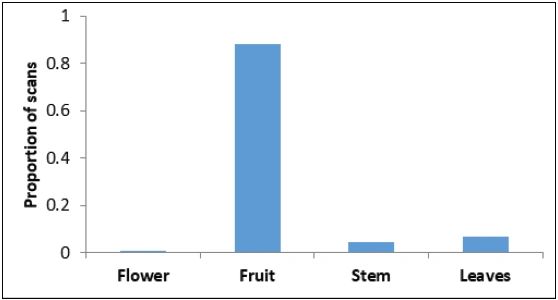
Figure 5: Proportion of different plant parts eaten by Nicobar long-tailed macaques
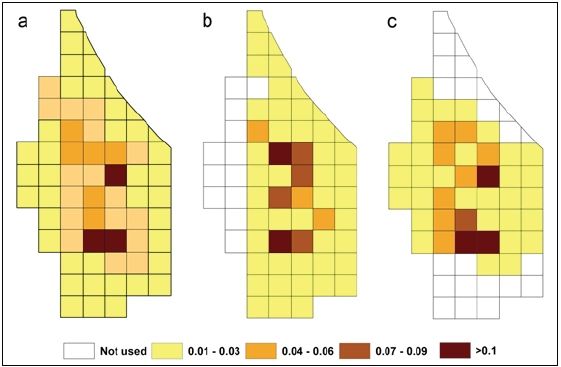
Figure 6: Proportion of grid cells used by Nicobar long tailed macaques in, a. Overall study period, b. Dry season, c. Wet seasonFigure
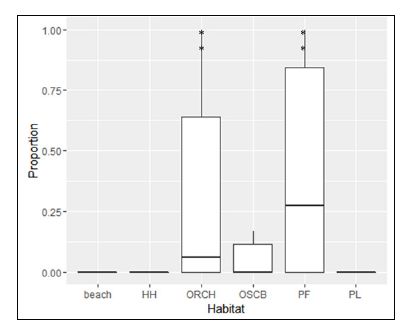
Figure 7: Proportion of habitat type in the grid cells within the home range of study group Nicobar long-tailed macaque. HH = Households, ORCH = Orchards, OSCB = Open Scrub, PF = Primary Forest, PL = Plantation. ** Tukey HSD test Padj<0.001
Table 2: Number and proportion of scans recorded for each activity in each season (total scans 4628)
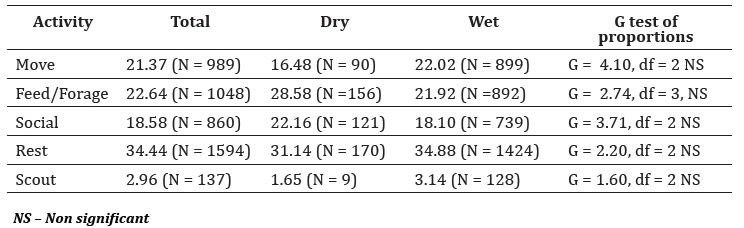
Table 3: Relative frequency and source of major food items consumed by Nicobar long tailed macaques.
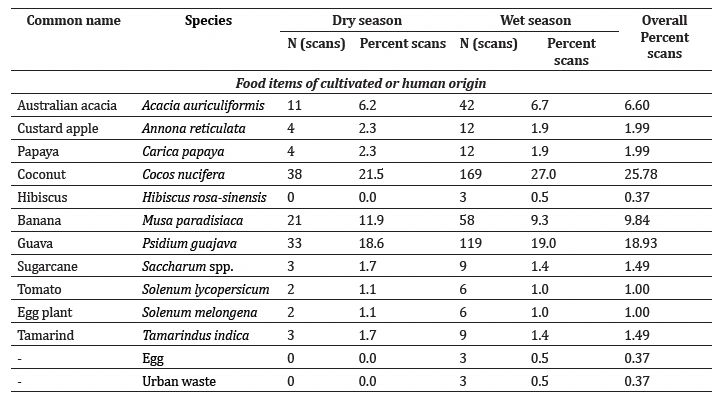
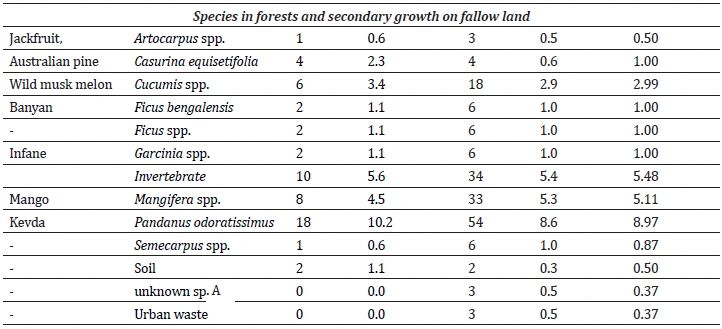
Table 4: Seasonal variation in daily path length in metres by Nicobar long-tailed macaques

Table 5: Influence of covariates on the intensity of grid use by Nicobar long-tailed macaques
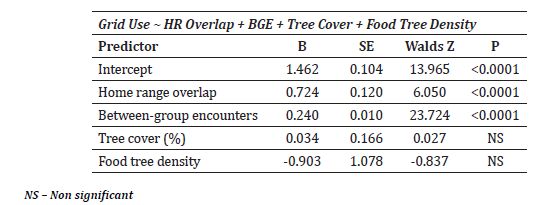
Table 6: Pearson’s product-moment correlation between the intensity of grid use and available habitat types.
The group was observed to feed on 25 food resources, of which four were most common, viz., coconut, banana, guava and Pandanus spp. Among them, three resources were from orchards and plantations in human habitation, and those were available throughout the year. Primary forests and orchards spatially cover the majority of the study area (72.2%). Yeager (1996) reported similar findings from Kalimantan Tengah, Indonesia, where the long-tailed macaque exploited 33 plant resources of which five were most frequent in their diet. This indicates the generalist nature of the species; as in both studies, macaques utilized the most abundant, and thus, low-cost resource first. Due to this high dependency on perennially available food sources, the group did not exhibit seasonality, and the most heavily used grids, or core home ranges, remained unchanged between the seasons. However, they were observed to utilize a few more grids, especially littoral patches, to feed on Pandanus, available during the dry season. Significantly longer daily path length in the wet season than in the dry season indicated a more intensive search of the area in the wet season.
The ranging pattern of a species is highly dependent on food availability, and seasonal changes in food availability (Clutton-Brock, 1975, Maruhashi et. al., 1998; Santosh et. al., 2015,). Human-influenced land cover types (orchards, plantations, households, and open scrub) constituted 58.6% of the home range of the present study group. This may have resulted in the constant availability of food resources in all seasons in the home range. Thus, the extent of intensively used grids by the study group did not differ significantly. However, the group used a larger portion of the home range in the dry season compared to the wet season. Such patterns have been observed in other macaque species also. In the toque macaques (M. sinica), the dry season home range of 3.98 ha was larger than the wet season range of 2.8 ha (Jayapali et. al., 2023). As in the case of the present study group which did not use many grids in the northern, western and southern parts of its range during the wet season, the Japanese macaques shifted their home ranges between the eastern and western parts and modified their travel rates seasonally (Hanya et. al., 2020). van Schaik et. al. (1983) demonstrated that larger groups travel farther due to the ‘pushing forward effect’ where group members try to go to newer unexploited patches sooner. The resource competition was very low in our study group, as it was a small unimale group. In a human-dominated landscape, the study group adapted to food resources available in human habitation, which has resulted in increased human-macaque conflict in the region. Coconut, banana, and guava are cultivated by local people for economic gains, and the resulting loss has increased human-macaque conflict (Mishra et. al. 2020c).
The between-group competition instead of food availability was the most influencing factor on grid-use by the study macaques. The study group was among the smallest compared to the two neighbouring groups of 79 and 37 individuals, having massive handicap during aggressive encounters (Pal et. al., 2018a). Hence, affinity of the study group towards home range overlap and areas with high between-group encounters seems apparently counter-intuitive. However, it can be explained due to the following reasons: van Schaik et. al. (1983) showed that daily path length increases with group size because individuals in a group attempt to reach intact foraging patches, and larger groups move often and spend less time in a patch. Conversely, a smaller group will exhaust the patch at a much slower rate and will not be influenced by ‘the pushing forward effect’. This may have allowed the present small group to utilize the smaller patches within the home range overlap more efficiently. Also, the resource requirement of a smaller group is lesser, enabling it to exploit smaller patches that larger groups cannot exploit. All these pattern show that the behaviour of the study group was typical of that of the commensal macaques of other species.
Acknowledgements
We thank the Department of Science and Technology, Government of India, for the financial assistance to HNK (Grant No.SR/SO/AS-49/2011 to carry out the project. We are grateful to the Principal Chief Conservator of Forests, Port Blair, Andaman and Nicobar Islands, for granting permission to carry out the study (Permit No. CWLW/WL/134/566), and the Assistant Commissioner, Andaman and Nicobar Administration, for providing the necessary permits to access the tribal areas. We express our gratitude to the Andaman and Nicobar Forest Department, especially the Nicobar division along with the Nicobar Administration, for logistical support. We thank the Directors, SACON, Coimbatore, for their valuable support. Mewa Singh thanks the Indian National Science Academy for the Distinguished Professor award (N0. SP/DP/2023/656) during which this article was prepared.
CONFLICT OF INTEREST
The authors have no competing interests to declare that are relevant to the content of this article.
DATA AVAILABILITY
Data and codes are available from the corresponding author on request.
AUTHOR CONTRIBUTIONS
HNK and ADV conceived the study. ADV, SVR, AP, and PSM carried out the fieldwork and data analysis. HNK and MS procured funding and supervised the study. All authors contributed to the article preparation.
Edited By
Vishnupriya Kolipakam
Wildlife Institute of India
*CORRESPONDENCE
Mewa Singh
✉ mewasinghltm@gmail.com
CITATION
Velankar, A. D., Kumara, H. N., Sheethal, V. R., Pal, A., Mishra, P. S. & Singh, M. (2024). Activity budget and habitat use in the Nicobar long-tailed macaque Macaca fascicularis umbrosus. Journal of Wildlife Science, 1(2), 87-95. https://doi.org/10.63033/JWLS.RUPT1872
COPYRIGHT
© 2024 Velankar, Kumara, Sheethal, Pal, Mishra, Singh, M. This is an open-access article, immediately and freely available to read, download, and share. The information contained in this article is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0![]()
![]() ), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepted academic practice. Copyright is retained by the author(s).
), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepted academic practice. Copyright is retained by the author(s).
PUBLISHED BY
Wildlife Institute of India, Dehradun, 248 001 INDIA
PUBLISHER'S NOTE
The Publisher, Journal of Wildlife Science or Editors cannot be held responsible for any errors or consequences arising from the use of the information contained in this article. All claims expressed in this article are solely those of the author(s) and do not necessarily represent those of their affiliated organisations or those of the publisher, the editors and the reviewers. Any product that may be evaluated or used in this article or claim made by its manufacturer is not guaranteed or endorsed by the publisher.

Edited By
Vishnupriya Kolipakam
Wildlife Institute of India
*CORRESPONDENCE
Mewa Singh
✉ mewasinghltm@gmail.com
CITATION
Velankar, A. D., Kumara, H. N., Sheethal, V. R., Pal, A., Mishra, P. S. & Singh, M. (2024). Activity budget and habitat use in the Nicobar long-tailed macaque Macaca fascicularis umbrosus. Journal of Wildlife Science, 1(2), 87-95. https://doi.org/10.63033/JWLS.RUPT1872
COPYRIGHT
© 2024 Velankar, Kumara, Sheethal, Pal, Mishra, Singh, M. This is an open-access article, immediately and freely available to read, download, and share. The information contained in this article is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0![]()
![]() ), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepted academic practice. Copyright is retained by the author(s).
), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepted academic practice. Copyright is retained by the author(s).
PUBLISHED BY
Wildlife Institute of India, Dehradun, 248 001 INDIA
PUBLISHER'S NOTE
The Publisher, Journal of Wildlife Science or Editors cannot be held responsible for any errors or consequences arising from the use of the information contained in this article. All claims expressed in this article are solely those of the author(s) and do not necessarily represent those of their affiliated organisations or those of the publisher, the editors and the reviewers. Any product that may be evaluated or used in this article or claim made by its manufacturer is not guaranteed or endorsed by the publisher.