TYPE: Research Article ![]()
Swati Nawani¹ , Deb Shankha Goswami², Arup Das³, Krishnendu Banerjee¹, Abhijit Das¹*
, Deb Shankha Goswami², Arup Das³, Krishnendu Banerjee¹, Abhijit Das¹*
¹Wildlife Institute of India, Chandrabani, Dehradun-248001, Uttarakhand, India.
²34, Talpukur south, Naihati, 24 Parganas(N), West Bengal, 743165, India.
³Aaranyak, 13 Tayab Ali Bylane, Beltola Tiniali, PO: Beltola, Guwahati – 781028, Assam, India.

Introduction
Stream amphibians, such as frogs, are valuable indicators of ecosystem health (Welsh & Ollivier, 1998). Nevertheless, they are vulnerable to climate change, deforestation, pollution, and habitat degradation (Kim et al. 2019; Panwar 2020). Frogs often exhibit site fidelity and limited home ranges, moving primarily within a small, defined area to access resources, avoid predators, and maintain body moisture and temperature (Sinsch 2014). Moreover, stream frogs inhabiting mountains at high elevations are more prone to habitat alteration and degradation than terrestrial species (Saeed et al. 2022). Thus, knowledge of how frogs use and move among habitats is crucial for effective management plans and strategies concerning the species and habitat (Groff et al. 2017). In recent years, radio telemetry has been used globally to study the movement, habitat use and home range of terrestrial and aquatic frogs (Daugherty & Sheldon, 1982; Tessier et al. 1991; Kam & Chen, 2000; Sinch et al. 2012; Akram et al. 2022). However, studies focused on the movement or home range of stream frogs in the Indian Himalayan region are unfortunately lacking, where biodiverse freshwater habitats are under threat (Johal & Rawal, 2005).
Herein, we investigated movement and home range of Nanorana vicina, using radio telemetry in India. This study aimed to address three questions, 1). Is there a seasonal difference in the movement of tagged frogs? 2). Are there differences in the movement pattern between a natural stream and another stream affected by anthropogenic disturbances? 3). What is the home range size of tagged frogs? We also assessed whether tags led to weight loss in frogs.
Materials and Methods
The study was conducted in Binog Wildlife Sanctuary (BWLS) (28° 60´ – 31°28´ N, 70°49´ – 80°60´ E), Uttarakhand, India (Figure 1). We selected two perennial streams and classified those as ‘Disturbed stream’ and ‘Undisturbed stream’ based on the degree of anthropological modifications such as check dams and artificial pools. Streams at the locality are spring-fed and approximately 5 meters wide with moderate flow, primarily rocky with intermittent gravel and pools with detritus, fallen logs and leaf litter. We used very high frequency external transmitters (Model SOPR- 2070, battery life 135 days; Wildlife Material, US), keeping in view the constrains in shape and size of the anuran body. Following global standards (Goldberg et al. 2002), a transmitter weighing less than 3% of the frog’s bodyweight was tied in the groin region. The frogs were kept overnight under controlled conditions before being released in the same area of capture. We tracked 16 individuals of N. vicina for an average span of 63 days (Minimum= 13 days; Maximum= 92 days) from May to October 2019 (Table 1). Bodyweight of individuals ranged from 115–190 gm (Mean± SD = 150.9 ± 24.58 gm, n = 16). In the initial month of the study, tagged frogs were located thrice at 7-8 AM, 12-1 PM, and 8-9 PM to assess diurnal and nocturnal movements. Subsequently, given the species’ nocturnal activity patterns, tracking frequency shifted to nightly readings between 8 PM and 12 AM, conducted near-daily to monitor potential movement. The surveys were mostly continuous, apart from a few sessions being abrupt by harsh weather conditions. Depending on the nature of the terrain the maximum range of the signal was 50 – 100 meters. We have used seven tags on seven individuals while four tags were reused on nine different individuals, which is explained in Nawani et al. (2022). Individuals were given identification Nanorana vicina, disturbed (NVD) and Nanorana vicina, undisturbed (NVU) followed by an individual identifying number. We used meter tape to calculate the daily distance moved by individuals after each relocation. We measured straight-line distances (from the previous location to the next location) between an individual’s movements. We collected location data with a focus on maintaining a five-meter range of accuracy. When a frog moved less than 10 meters, we decided not to record the location as GPS errors could potentially compromise the reliability of such data. However, when we could achieve accuracy levels better than the individual’s movement range, we recorded GPS locations (average = 13 locations per individual, n = 6). Consequently, individuals (n = 10) with very less movement or location data were excluded from home range estimation. We discontinued tracking in case of transmitter signal failure or a situation where the transmitter began to abrade any frogs.
Data Analysis:
Since the sex of the individuals (male and female) could not be ascertained based on their morphological characteristics in the field, we refrained from including any tests or analyses related to this aspect. Using the movement data, we calculated the mean distance moved by all individuals over the study period. To determine whether there were differences in the seasonal movement, we performed Kruskal-Wallis test with pre-monsoon (May-June), monsoon (July–August) and post-monsoon (September–October) movement data, followed by Dunn’s test with Benjamini-Hochberg adjustment. We also calculated the percentage of total movement made in streams and land. We performed the Mann-Whitney U test to compare movements made in two streams. Our sample size was low; therefore, we calculated home range using minimum convex polygon (MCP) at 50% and 90% isopleths. We also assessed the change in the weight of frogs before tagging and after the final removal of tags using the Wilcoxon’s signed rank test. The values given are mean with standard error and the significance for all tests was compared at p-value = 0.05. We performed statistical analysis using Microsoft Excel and R version 3.4.1 (R core team ,2021). All maps were created using Arc GIS version 10.1 (ESRI, Redlands, USA).
Results
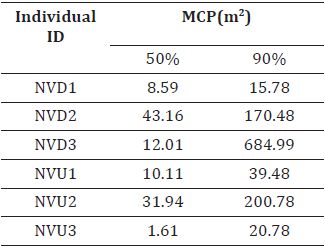
During the study period, the majority of frog movement (90.60%) was confined to the wetted width of the stream, with only 9.39% occurring on land. The mean movement distance varied among individuals, ranging from 0.49 ± 0.14 meters to 5.9 ± 3.26 meters (Table 1). Individual home ranges, estimated using the MCP method, exhibited substantial variation. At the 50% utilization level, home ranges spanned 1.61 m² to 43.16 m², while at the 90% utilization level, they encompassed a much wider area of 15.78 m² to 684.99 m² (Table 2).
Frog movement exhibited significant seasonal differences (df = 2, p << 0.05). Movement was the highest during the monsoon season (2.71 ± 0.46 m), followed by the pre-monsoon (1.41 ± 0.12 m) and post-monsoon seasons (0.78 ± 0.14 m). Post-hoc analysis using Dunn’s test revealed that while pre- monsoon and monsoon movement did not differ significantly (p > 0.05), post-monsoon movement was significantly lower than both pre-monsoon (p << 0.05) and monsoon (p << 0.05). No significant differences were observed in movement distances between frogs tagged in the two different streams (p > 0.05).
Furthermore, no significant differences were found in the body weight of the individuals before tagging and after removing the tags (n = 10, p > 0.05)
Discussion
Previous studies on N. vicina movement patterns have yielded valuable insights, albeit with certain limitations. Akram et al. (2022) observed restricted movements of less than 3 meters during a short-term, post-monsoon survey (8 days), while Batool et al. (2023) reported movement within a 120 m² area along stream banks. These studies, while providing valuable insights, primarily focus on short duration observations and do not offer a comprehensive assessment of home range. Our study, employing the MCP method, offers a broader understanding of the home range of N. vicina. Furthermore, our study covers three distinct seasons (pre-monsoon, monsoon, and post-monsoon), capturing potential seasonal variations in movement.
MCP analysis revealed that the majority of movements by tagged individuals in our study were concentrated along the stream course, with occasional movement towards land (Figure 2). This pattern aligns with previous studies and highlights the species’ close association with stream habitats. The use of MCP in our study is justified, as it is the standard method in studying home range in frogs (Luger et al. 2009; Neu et al. 2016) and can be used when the sample size is low (Boyle et al. 2009). While the sample size may limit the generalizability of our findings, our results, combined with those of Akram et al. (2022) and Batool et al. (2023), collectively suggest that N. vicina exhibits a restricted movement pattern, particularly in association with stream environments. This understanding is crucial for conservation efforts, as it highlights the species’ vulnerability to habitat alterations, particularly those affecting stream ecosystems.
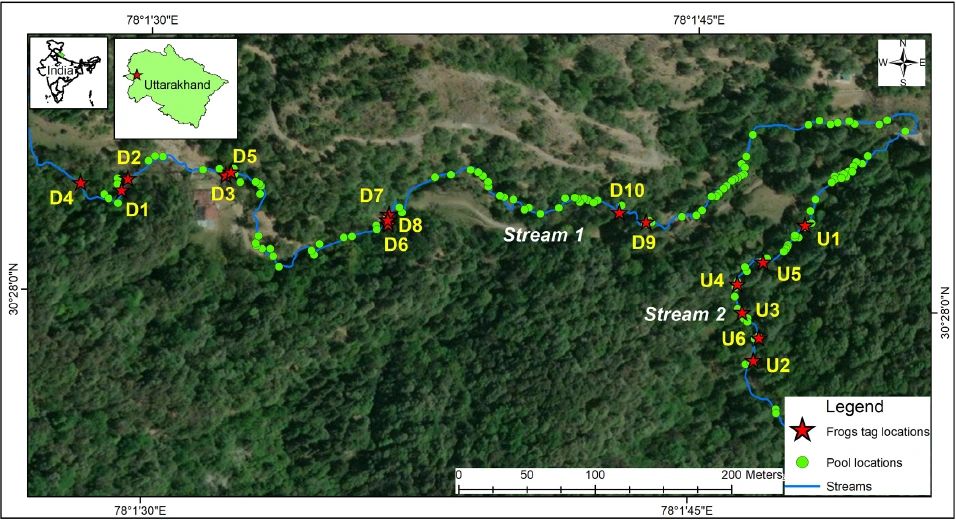
Figure 1: Locations of the tagged Nanorana vicina, movements in Binog Wildlife Sanctuary, Uttarakhand, India. (D1 to D10 are the individuals in the disturbed stream (n=10), and U1 to U6 are the individuals in the undisturbed stream (n=6).
Table 1. Movement and tracking history of 16 radio tagged frogs.
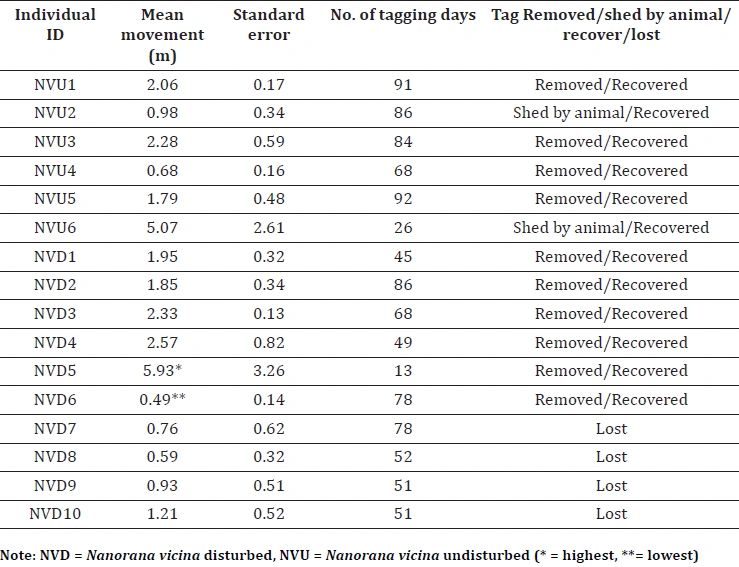

Figure 2. The home ranges (MCP) of six radio- tagged Nanorana vicina, individuals along the streams and riparian area. The width of the streams is approximately 5 m. (A) The home ranges in the disturbed stream, (B) the home ranges in the undisturbed stream.
Table 2. Home range at 50% and 90% of six individuals using Minimum convex polygon.
Although frogs exhibited less vagility, tagged frogs moved slightly more during the monsoon. In one case, a single individual (NVD3) moved approximately 100 m in a day during the monsoon (July), which was the highest movement throughout the study period. This movement resulted in the largest home range for NVD3 individual (Figure 2, A). Studies have shown that such movements by species may be undertaken to avoid areas that could flood during rain (Bosman et al. 1996).
While our study results do not provide an explanation for factors influencing movement and philopatry in N. vicina, the species’ affinity for pool habitats with rocky boulders, as noted by Ahmed et al. (2020) and Rais et al. (2014), may offer a partial explanation. The abundance of such habitats in both streams could explain the lack of significant differences in the observed movement patterns between them. Additionally, the larval morphology of N. vicina, characterized by an anteroventral oral disc, dorsoventrally flattened body, and low tail fins, suggests adaptation to shallow, flowing water environments (Banerjee et al. 2020; Gill et al. 2020). A similar pattern has also been reported in some tropical and temperate stream-dwelling anurans that exhibited philopatry with limited movement (Inger 1969; Daugherty & Sheldon, 1982; Tessier et al. 1991). The limited movement by individuals is probably due to easy accessibility of necessary resources in their nearby surroundings, such as riparian zones. These areas probably provide sufficient ecological resources, therefore long-distance movements are not required (Kam & Chen 2000) and amphibians tend to move less during their lifetime compared to the other vertebrates (Zug et al. 2001). While our study did not find significant differences in overall movement between disturbed and undisturbed streams, previous work in the same study area has documented the impact of check dams and stream channel modification on tadpoles of the genus Nanorana (Jithin et al. 2022a, b; Jithin & Das 2022).
Additionally, the tag attachment did not significantly impact the body weight of the frog. Tags were immediately removed if any lesions on the body of the frogs were observed, ensuring animal welfare. Furthermore, two frogs with attached nonfunctional tags were recovered after a span of 21 months;, however, no weight loss was detected. Throughout the course of study, we successfully recovered seven tags from the individuals while the rest four tags were lost (Nawani et al. 2022).
Our results provide valuable insights into the movement and home range of this sedentary, stream-dwelling frog. Thus, serving as a foundational platform for future research on this specific species and other Himalayan stream frogs. However, further studies are required to answer the factors influencing the movement, habitat use and home range of stream frogs. Additionally, to comprehend the impacts of habitat degradation on these less mobile and sedentary species, a long-term monitoring study is required.
Study Limitations:
While our study add new information about the movement patterns and home range of N. vicina, certain limitations need to be acknowledged. The species’ limited movement, particularly if less than 5 meters, coupled with the GPS device’s location error of 5 meters, may have compromised exact home range estimation. To mitigate this potential bias, we recorded GPS locations only when frogs moved more than 10 meters and supplemented these data with manual measurements using a meter tape.
Additionally, our home range analysis was based on six individuals. Thus, low sample size may limit the generalizability of our findings to the broader population of N. vicina. Further studies incorporating larger sample sizes and potentially more precise tracking technologies would be worthwhile to refine our understanding of N. vicina home range dynamics.
Acknowledgement
We sincerely acknowledge – Uttarakhand Forest Department providing research permission (permit letter no 1236/5-6 [WII] dated 16 November 2017). We also acknowledge Director, Dean and Research Coordinator of WII and Indraneil Das (Malaysia) for supporting our research. We are extremely grateful to Mrs. Kehkashan Naseem (Former DFO) and Dr. Shipra Sharma (Former FRO) for their immense support during field work. We would also like to extend our thanks to Forest guards of Binog wildlife sanctuary. Authors are thankful to Nilanjan Chatterjee, Jithin Vijayan, Siddhart Rajmohan, Shruti Arora, Aastha Saini, Naitik Patel, Bitupan Boruah, Anukul Nath, Bhawna Dhawan, Surojit Moitra, Devendra Ji and his family, Kuldeep Rawat and his family. We would like to express our sincere gratitude to Mr. Varun S Kher, Scientist C, WII and the anonymous reviewer for their time and expertise in reviewing our manuscript.
CONFLICT OF INTEREST
The authors have no competing interests to declare that are relevant to the content of this article.
DATA AVAILABILITY
The data that supports the findings of this study are available in this article..
AUTHOR CONTRIBUTIONS
Conceptualization, fundraising and editing: Ab.D,
Field Work and data collection: S.N, D.S.G, K.B, Ab.D,
Data analysis: S.N, Ar.D, K.B.,
Original draft writing: S.N, K.B.
Edited By
Mewa Singh
Institution of Excellence, University of Mysore
*CORRESPONDENCE
Abhijit Das
✉ abhijit@wii.gov.in
CITATION
Nawani, S., Goswami, D. S., Das, A., Banerjee, K., Das, A. (2024). Home range and seasonal movement of Nanorana vicina, (Stoliczka, 1872) (Anura: Dicroglossidae) in Himalayan streams, India. Journal of Wildlife Science,1 (2), 81-86. https://doi.org/10.63033/JWLS.JJAP3927
FUNDING
Funding was supported under the Grant in Aid scheme from the Wildlife Institute of India (WII), Dehradun and the Ministry of Environment, Forest and Climate Change, Government of India.
COPYRIGHT
© 2024 Nawani, Goswami, Das, Banerjee, Das. This is an open-access article, immediately and freely available to read, download, and share. The information contained in this article is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0![]()
![]() ), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepted academic practice. Copyright is retained by the author(s).
), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepted academic practice. Copyright is retained by the author(s).
PUBLISHED BY
Wildlife Institute of India, Dehradun, 248 001 INDIA
PUBLISHER'S NOTE
The Publisher, Journal of Wildlife Science or Editors cannot be held responsible for any errors or consequences arising from the use of the information contained in this article. All claims expressed in this article are solely those of the author(s) and do not necessarily represent those of their affiliated organisations or those of the publisher, the editors and the reviewers. Any product that may be evaluated or used in this article or claim made by its manufacturer is not guaranteed or endorsed by the publisher.

Edited By
Mewa Singh
Institution of Excellence, University of Mysore
*CORRESPONDENCE
Abhijit Das
✉ abhijit@wii.gov.in
CITATION
Nawani, S., Goswami, D. S., Das, A., Banerjee, K., Das, A. (2024). Home range and seasonal movement of Nanorana vicina, (Stoliczka, 1872) (Anura: Dicroglossidae) in Himalayan streams, India. Journal of Wildlife Science,1 (2), 81-86. https://doi.org/10.63033/JWLS.JJAP3927
FUNDING
Funding was supported under the Grant in Aid scheme from the Wildlife Institute of India (WII), Dehradun and the Ministry of Environment, Forest and Climate Change, Government of India.
COPYRIGHT
© 2024 Nawani, Goswami, Das, Banerjee, Das. This is an open-access article, immediately and freely available to read, download, and share. The information contained in this article is distributed under the terms of the Creative Commons Attribution License (CC BY 4.0![]()
![]() ), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepted academic practice. Copyright is retained by the author(s).
), allowing for unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited in accordance with accepted academic practice. Copyright is retained by the author(s).
PUBLISHED BY
Wildlife Institute of India, Dehradun, 248 001 INDIA
PUBLISHER'S NOTE
The Publisher, Journal of Wildlife Science or Editors cannot be held responsible for any errors or consequences arising from the use of the information contained in this article. All claims expressed in this article are solely those of the author(s) and do not necessarily represent those of their affiliated organisations or those of the publisher, the editors and the reviewers. Any product that may be evaluated or used in this article or claim made by its manufacturer is not guaranteed or endorsed by the publisher.